There is no record of the number of people who attended the Open Day for bird ringing at Grotto Bay, but it was probably roughly 40, both residents and visitors.But we do know that 76 birds of 27 species were handled. There were eight retraps, so the number of birds ringed was 68! The list is near the bottom of this report. This was our third Open Day to the Grotto Bay Private Nature Reserve, and reports on the two previous visits are here (27 May 2023) and here (31 August last year).
Fortunately, in spite of dire and persistent predictions to the contrary, the wind was missing in action. Mist nets hanging like curtains in one of the firebreaks at Grotto Bay.
The wind clearly did not read the forecast that it was supposed to be doing 27 to 30 km per hour in the critical period 05h00 to 08h00, with gusts of 38 to 45 km per hour. It failed to get up and stayed in bed. And for that we were grateful.
The veranda of the Grotto Bay Conservation Centre is a marvellous resource for bird ringing, and for explaining bird ringing.
The Conservation Centre at Grotto Bay is a few hundred metres farther inland from the village itself. Just in front of it is a small wetland. This is the view from the veranda. It was a high-quality day, both weatherwise and action-wise!
Extracting a bird safely and gently from a mist net takes real skill. This white-irised bird is a Layard’s Warbler.
We caught birds at a steady pace, but were never overwhelmed.
The wonderful thing about bird ringing is the opportunity to see birds up close.
This is a young Bokmakierie
Head of Karoo Prinia.
Head of a juvenile Long-billed Crombec.
Adult male Southern Masked Weaver being ringed. The leg is held firmly but gently at the “knee” during this operation, so that it can’t move.
The weaver took advantage of a moment’s lapse in concentration of the ringer! The beak is strong – it weaves nests – and although it can deliver a good pinch, it does not draw blood.
The marie biscuit-coloured eye indicates that this is an adult male Cape Weaver.
Measuring the “total-head length” of a juvenile Yellow Canary with dial calipers. The reading is 25.6 mm. The 20 comes from the yellow part of the calipers. The dial makes one revolution in 10 mm. It is at 5.6 mm. So 20 + 5.6 is 25.6 mm.
There is nothing quite like bird ringing to help develop connectedness to nature.
Daniel is using the gadget which helps us decide on the appropriate ring size for the Malachite Sunbird he has in the ringer’s grip. There will be stories to tell at school on Monday morning.
The fieldguide is open at the Cape Weaver page. This is a remarkable way to learn the identification features of a bird!
Hybrid bulbul?
The morning was invaluable from the social science perspective, in achieving its Open Day objectives! From the natural science perspective, we helped expand the SAFRING database. But the stand-out and intriguing piece of natural science was a bulbul which is possibly (maybe even probably) a hybrid between a Cape Bulbul and an African Red-eyed Bulbul.
Both of these birds had brood patches, and both were adults. The bulbul on the left is Cape Bulbul. We suspect the bird on the right is a hybrid between a Cape Bulbul and an African Red-eyed Bulbul. In the Cape Bulbul, the white cere around the eye is an ellipse, and the eye is at the back end of the ellipse. In this bird, the short axis is 65% of the length of the long axis. For the bird on the right, the short axis is 88% of the length of the long axis, and the eye is almost in the centre of the cere. In African Red-eyed Bulbuls, illustrated below, the cere is bright red, and circular. The colour of the cere of the suspected hybrid is a bit of a mess, a kind of pink intermediate between white and red. The fact that the cere of the bird on the right is more circular than the norm for a Cape Bulbul also adds fuel to the idea that it is a hybrid.
This is an African Red-eyed Bulbul. Chief differences with Cape Bulbul are the circular red cere, the darker head, and the whiter belly.
The suspected hybrid, on the right, has a slightly whiter belly than the Cape Bulbul on the left. Ringers have picked up other examples of suspected hybrids recently. In the northern part of the Western Cape, in the Olifants River valley as far south Citrusdal, most of the bulbuls are now African Red-eyed. There are even records of African Red-eyed Bulbul at Stanford and even at Cape Agulhas; see this paper in Biodiversity Observations.
Here is the list of the 27 species at Grotto Bay, and the number of birds of each species handled.
There is a list of all past BDI ringing events that have reports here. The next ringing event is also an Open Day. It is in Saturday 20 December, with ringing on the western side of Zandvlei, followed by a bring and brunch at Marina da Gama, on the eastern side. Details here. Information about future ringing plans are here.
Thank you
First and foremost, all the visitors want to thank the Grotto Bay Private Nature Reserve for the warm welcome, and the generous hospitality. The muffins, the shortbread, the quiches, …, …, … were amazing and greatly appreciated. Especial thanks to Roy Lawrence, Michelle van Acker, Suzette Vorster, and also to Loutjie Steenberg, who made the original arrangements, but in the end suffered FOMO in being unable to attend. Many of the photographs in this blog are Suzette’s – thank you.
Ringers, plus family and friends, trainees and would-be trainees are invited to the end-of-year event on Saturday 20 December 2025. We will be bird ringing at Zandvlei, then have brunch at Marina da Gama.
We inaugurated this last year, and the report on the event is here.
Networking after the ringing last year!
We will set up a WhatsApp group to communicate final details.
Sunrise is at 05h31 that day. So the ringing will be starting early! Details of times and place(s) will be sent to everyone who responds. You don’t need to come as early as the start of the ringing, and you are even welcome to just come for the brunch! We will set up a WhatsApp group to communicate final details.
Brunch will be a bring and share event. Like it was last year:
In spite of the appearances here, there were lots of people to tackle all the food. Last year, this included an ostrich egg omelette!
This event provides a great opportunity to network, and to make plans for ringing in 2026.
Please contact Sue Gie if you would like to be part of the morning; send an email to susan[at]gie.co.za
The BDI ringing programme for 2026 is here. There is a list of all the events which have reports here.
Bird ringing has made a huge contribution to bird research and bird conservation. This is outlined here.
We have been monitoring the breeding of the African Oystercatchers Haematopos moquini on Robben Island since the summer of 2001/02. We have also monitored their abundance. This blog aims to describe the fieldwork we are doing in the summer of 2025/26. Our target interval between trips is six days, but that sometimes gets adjusted by the weather and other commitments. So this blog ought to be updated at roughly six-day intervals. The most recent visit is on top! Our objective for the summer is to find every oystercatcher nest on the island. At the end of the blog is a list of some of the publications that have emerged from this fieldwork. The papers give more background to the science and conservation objectives.
Fieldwork 10 : 16 and 17 January (in progress)
This fieldtrip had been scheduled for the 15th and 16th. But the sea was unsuitable for operation on the 15th, so we went to the island with the first ferry on the 16th, had a long day’s fieldwork, and continued into the morning of the 17th. It now feels like we have caught up!
This is our first nest actually in Murray’s Bay Harbour. The nest owner is watching from along the edge of the wharf!
This is probably the first nest that we have found on the inland side of the perimeter road. This is a busy spot with Alpha One in the background. Every tourist bus passes this spot twice. The nest has had two eggs since it was discovered on 5 January. So it is about one-third of the way through the 30-day incubation period.
Fieldwork 9 : 10 January 2026 (in progress)
The two-day trip which had been planned for 3 to 4 January was unable to take place because of high winds and no ferries. As luck would have it, it proved impossible to arrange fieldwork before the next planned dates of 9 and 10 January. And then ferries were cancelled on 9 January, and fieldwork was reduced to a day trip on 10 January.
Fieldwork 8 : 27 December 2025 (in progress)
This was a planned one day trip, due to it being in the holiday season.
Dembo Jatta assisted with almost all of the fieldwork up to this trip, and was the leader of fieldwork in the northern half of the island. This was his last trip before starting a PhD at Wageningen University in the Netherlands; this is the opportunity to thank him for his dedication to the oystercatcher monitoring this season, and to wish him success with the next phase of his studies.
Fieldwork 7 : 21 and 22 December
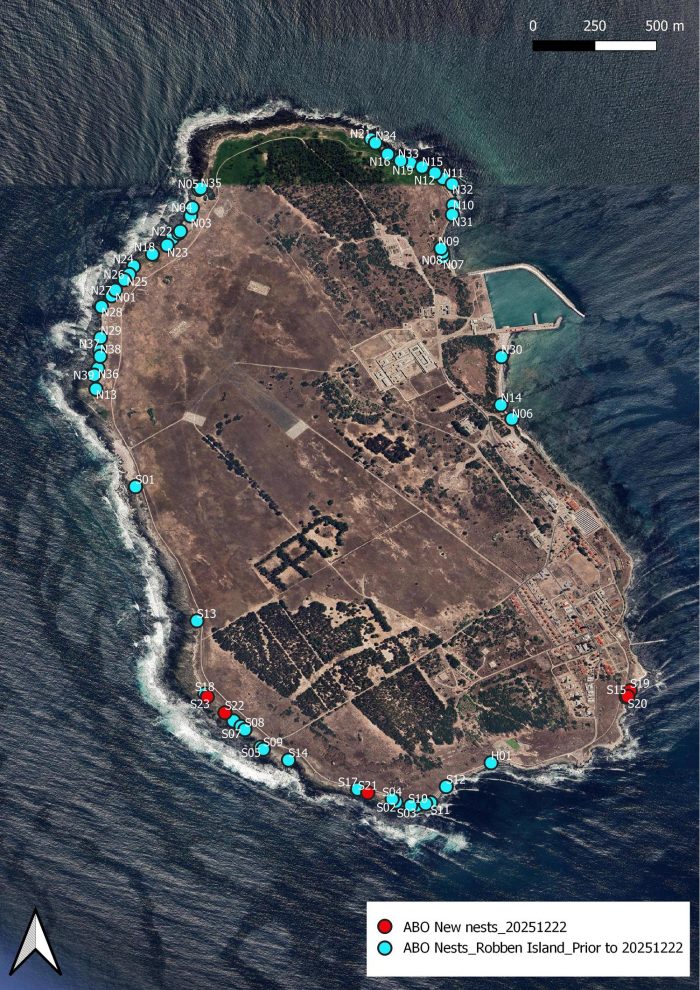
This is a report on the 7th fieldwork trip of the 2025/26 oystercatcher breeding season. We found six new nests with eggs (five in the southern half, nests S19 to S23; and one in the northern half, N40). This brings the total number of nests found to 63.
When you see the eggs, they are obvious. Nest S21.
Both of the chicks that were ringed during Fieldwork 5, on 8 and 9 December, are doing fine. The parents no longer bother to hide them, because they can fly. This chick is 42 days old; they fly at about 40 days old. It still has lots of growing to do.
The rusty ship’s boiler on the shore marks the end of the afternoon’s fieldwork on Sunday at 19h00. The team will be back here at 06h30 to continue.
The new nests are in red. The position of nest N40 still needs to be added. It lies between N14 and N30, just south of the harbour.
Bird list
This is the bird species list for Fieldwork 7. Two species, in bold, were recorded for the first time this summer. The list has 38 species.
Species
Note
Cisticola, Zitting
Along the road between the foghorn and the airfield
Cormorant, Bank
Cormorant, Cape
Cormorant, Crowned
Cormorant, White-breasted
Only seen at the harbour; maybe three present.
Crane, Blue
Over airfield, harassed by Kelp Giull
Crow, Pied
Dove, Red-eyed
Dove, Ring-necked
Egret, Cattle
Egret, Little
Fiscal, Southern
Goose, Egyptian
Guineafowl, Helmeted
Gull, Hartlaub’s
Gull, Kelp
Ibis, African Sacred
Ibis, Hadada
Kestrel, Rock
One seen, hunting from tree
Night Heron, Black-crowned
One, just after dawn apparently flying from the coast to its daytime roost
Oystercatcher, African
We have our first fledglings!
Partridge, Chukar
Penguin, African
Pigeon, Speckled
Plover, White-fronted
Sparrow, House
Seen in three places! Harbour, research house, in the village
Spurfowl, Cape
Starling, Common
Swallow, Barn
Swallow, White-throated
Swift, Little
Tern, Common
Tern, Greater Crested
Tern, Sandwich
Thick-knee, Spotted
Turnstone, Ruddy
One, south of Bluestone Quarry
Weaver, Cape
Whimbrel, Eurasian
Total
38 species
We made the second ever record for the Robben Island of Blue Crane. It was circling above the northwestern corner of the island, over the remains of the airfield, which dates back to the Second World War. This northwestern part of the island is part of the scattered Kelp Gull breeding colony, so it is not a surprise that the intruder was mobbed by a Kelp Gull. See the photos below.
The most recently added species was African Dusky Flycatcher, on 23 May 2024. See the paper First record of African Dusky Flycatcher on Robben Island published in Biodiversity Observations.The total bird list for Robben Island remains on 166, but the list of species recorded since 2000 increases to 136. The most useful paper on Robben Island’s birds is available here. It was published way in back in 2011, so it needs an update!
Fieldwork 6 : 15 and 16 December
This is a report on the 6th fieldwork trip of the 2025/26 oystercatcher breeding season. We found 14 new nests with eggs (four in the southern half, nests S15 to S18; 10 in the northern half, N30 to N39). This brings the total number of nests found to 57. See the map below. We found our first three-egg clutch of the season. Here it is … nest N37:
37 of the 57 nests found with eggs still have eggs. Probably 10 pairs of the 57 nests have chicks which are alive, judged by the fact that the adults are alarming when we pass through their territories. So the are about 10 pairs which either lost eggs or small chicks. It is still early enough in the breeding season for these pairs to produce a replacement clutch. These are produced on average about 14 days after the loss of eggs or chicks.
Territories are short sections of the shore, and so nests are close together, so we can’t always be certain which chicks originally belong to which nests. But this chick probably comes from N01; from the records, it probably hatched on about 20 November, so on 15 December it would have been about 25 days old. It was being dive-bombed by Kelp Gulls.
The African Oystercatchers seem to have had a particularly good breeding year so far. Relatively few nests have been lost. Eggs which were laid before the middle of November have hatched by now (incubation period of about 28 days) so there are chicks at all stages of development, from hatchlings onwards. Chicks fly at around 40 days, at about two-thirds of adult weight.
This is the distribution of nests around Robben Island. The newly found nests are in red. Nests fund during Fieldworks 1 to 5 are in blue.
As on previous rounds of fieldwork, we found lots of scrapes. The scrape is placed alongside strands of dry kelp. When the oystercatcher is incubating, it will not be conspicuous! Hopefully many of these scrapes will become red dots on the map, with eggs found in them soon …
This scrape is on the southern half of the island. There are lots of territories here. Currently, there are only 18 nests in the south, compared with 39 in the north!
Birds
This is our bird list, in alphabetical order. The six species in bold were seen during Fieldwork 6 for the first time this summer.
Species
Comment
African Oystercatcher
We are on 57 nests so far.
African Penguin
Several with penguin cellulitis were observed (see photos below) and reported. They have since been collected and admitted to SANCCOB for treatment.
African Sacred Ibis
Bank Cormorant
Blacksmith Lapwing
Cape Cormorant
Cape Spurfowl
Caspian Tern
One feeding along shore north of harbour
Chukar Partridge
Common, with chicks
Common Starling
Crowned Lapwing
The chicks of the pair on the football field are large
Egyptian Goose
Isolated birds along the shore, and three in Van Riebeek’s Quarry
Eurasian Whimbrel
Fiery-necked Nightjar
One heard along Boundary Road in evening
Fiscal Flycatcher
One seem along Boundary Road
Greater Crested Tern
Gathering. Pairs starting to do display flights
Hadada Ibis
Hartlaub’s Gull
About 30 at harbour
Helmeted Guineafowl
House Sparrow
One seen at harbour, one at research house
Kelp Gull
Breeding around north end of island, from the harbour to just north of Boundary Road. Lots of chicks of assorted sizes. Fledged young.
Little Egret
Little Swift
Pied Crow
At least five on the island
Red-eyed Dove
Ring-necked Dove
Rock Kestrel
One on tall tree
Ruddy Turnstone
Sandwich Tern
One at harbour
Southern Fiscal
Speckled Pigeon
Spotted Thick-knee
Western Cattle Egret
Small numbers
White-throated Swallow
White-breasted Cormorant
White-fronted Plover
Yellow-billed Kite
One seen circling over central grassland
… the right eye and the left eye of the same Speckled Pigeon. Non-circular pupils!
African Penguins have more than enough problems to deal with. Here is a new one. The bird in the centre of the photograph below has a newly occurring condition which has been dubbed cellulitis. Look at the fat legs. The beautiful sleek feathers indicate that it has spent a couple of weeks on the shore doing moult. It is now desperately needing to get back to the ocean to regain condition, but it has accumulated fluid in the legs. The cause of the condition is unknown. We spotted several penguins like this, provided exact coordinates, and they have been admitted to SANCCOB, where the skills exist to treat this condition:
The middle two penguins below are serious cases of penguin cellulitis. The penguin at the back is still moulting.
The feather replacement phase of moult takes about 14 days. They come ashore looking like garden gnomes. Even the flippers are full of fat. They are on land for about three weeks living off the fat reserves, sustaining not only themselves, but doing the energetically process of growing a complete new coat of feathers simulataneously. The annual moult is a mega-challenge for penguins. They can’t skip moult, because the old feathers lose their ability to enable the penguin to swim in a bubble of air, and keep the cold out.
This was our first Caspian Tern of the summer. It was feeding over the shoreline north of the harbour.
Egyptian Geese in Van Riebeek’s Quarry, south end of the island.
Fieldwork 5 : 8 and 9 December 2025
This view is always a reminder of the privilege it is to do research on the Robben Island World Heritage Site. We are grateful to the Robben Island Museum for the opportunity to do fieldwork here, and for the logistics provided.
On this fifth fieldwork trip of the 2025/26 African Oystercatcher breeding season, we found 13 new nests (S14 in the southern half of the island, and N19 to N29 in the northern half). The northern half is racing ahead with twice as many nests as the southern half. The total number of nests found at the egg stage is now 43. None of the clutches so far have had three eggs.
We found one dead ringed oystercatcher: ring K50080. The SAFRING database says that it was ringed on 5 February 2018 as an adult male.
We ringed the two chicks which hatched on 11 November 2025 on Trip 1. See far below! They have grown from …
… this fluffball size, covered in down, to …
… this almost completely feathered chick is 29 days old. All oystercatcher chicks weigh close to 39 g when they hatch. When we ringed this one, it weighed 421 g and the other chick was 355 g. When there are two chicks, the weights often diverge as they get older. They are capable of flight at around 470 g, which is about two-thirds of adult mass. They are still growing when they start flying!
Chicks are starting to hatch in quite a few of the first-laid nests. For example, here is N17 …
Our egg measurements and the masses when we found nest N17, and our formulae, suggested that incubation for these two eggs in N17 started on 8 November. The average incubation period is 30 days; this takes us to a stab at hatch date of 8 December. These two fluffballs were observed, still in the nest, on 9 December, so our method works well (at least in this case!). The ice cream tub is ours, but the immediate surroundings of this nest contain an alarming amount of little pieces of plastic and other rubbish. The litter does not come from the island; much of it gets dumped into Table Bay by the Black River which is the stormwater drain for much of the City of Cape Town. If you didn’t read this blog as part of Fieldwork 2, read it now: Industrial biodiversity 2 : Black River in Paarden Island : stormwater drain.
If you failed to spot the N17 fluffballs in the photo above, here is a second opportunity, and a chance for a close up view of a piece of litter.
This egg is “starred”. The hatchling will emerge, but the process of getting out of the shell is measured in days, not hours!
This is the state of play after Fieldwork 5. The nests found this visit are shown in red. They are strikingly clustered. The positions of nests found during the earlier fieldwork sessions are in blue. Diane Smith produced this map.
Bird list
Bird list in alphabetical order. It is our second longest list so far this summer, at 36 species. The longest had 41 species, during Fieldwork 1.
Species
Comment
African Oystercatcher
Breeding deeply underway (see above!)
African Penguin
Small numbers moulting
African Sacred Ibis
Flying to and from mainland
Bank Cormorant
Breeding at short arm
Barn Swallow
One seen hawking over shore
Blacksmith Lapwing
A pair in the Blue Stone Quarry
Black-crowned Night Heron
One on the shore close to the Sea Challenger, late afternoon
Black-winged Stilt
2 at Bluestone Quarry; third record for the island (see below)
Cape Batis
One in “woodland” alongside Boundary Road
Cape Cormorant
Cape Spurfowl
Groups include young birds
Cape Weaver
Chukar Partridge
Groups include young birds
Common Starling
Common Tern
One at harbour
Crowned Cormorant
Crowned Lapwing
Pair with young on football field
Egyptian Goose
A few at the Blue Stone Quarry
Eurasian Whimbrel
Several
Great Cormorant
One at harbour
Greater Crested Tern
Hundreds gathering
Hadada Ibis
A few
Hartlaub’s Gull
Mostly at the harbour
Helmeted Guineafowl
Groups include young birds
Kelp Gull
Breeding; both eggs and young
Little Egret
A handful, scattered along coast at rockpools
Pied Crow
Scattered. Possibly five
Red-eyed Dove
Ring-necked Dove
Ruddy Turnstone
Small group
Southern Fiscal
Scattered across island; juveniles present
Speckled Pigeon
Lots
Spotted Thick-knee
Heard at night near research house
Western Cattle Egret
Arrive from mainland in evening
White-fronted Plover
Second nest found
White-throated Swallow
A pair at the harbour
There were at least two Black-winged Stilt at the Bluestone Quarry. This is the third record for the species on Robben Island. The first was made September and December 2012 (see First observation of Black-winged Stilt Himantopus himantopus on Robben Island), and the second was made between November and December 2017, on checklists for the Second Southern African Bird Atlas Project. Unfortunately, there are no photographs, but hopefully the stilts will stay for a while, as on the previous two occasions, and we will get a photographic record during Fieldwork 6!
The tips of the outermost primaries of this Common Tern are starting to disintegrate. They will likely have something like 30,000 km on the clock since they were replaced in around February-March this year. A northwards and a southwards migration, and lots of flying while on the breeding grounds. Bird ringing shows that most of the Common Terns that migrate to South Africa breed in the countries around the Baltic Sea: especially Finland and Sweden.
White-fronted Plover.
Mole snake
Mole Snake, c. 40 cm, young specimen still showing some white spots. This was the only Mole Snake seen during Fieldwork 5.
Fallow deer
Here is a young fallow deer boldly walking along the main road between the village and the harbour on Robben Island, at 09h00 on a week day morning!
Stop the Fallow Deer. There is a tiny number of adults left. But if left, it’ll grow rapidly again to another large herd. They are destructive to the vegetation, which is recovering amazingly after the ravages of centuries of rabbits.
Plastic
Besides the plastic in and around the nest above, here is a plastic bottle …
Dianne found this water bottle on Robben Island. Internet archaeology revealed that Nongfu Spring is the top supplier of bottled water in China. You are invited to visualize the journey this plastic took from source to shore.
Nightlife
The City of Cape Town, from Robben Island, 8 December 2012. Table Mountain is floodlit.
Fieldwork 4 : 3 and 4 December 2025
We found six new nests (S13 on southern half, and N14 to N18 in the north) . Total nests found with eggs so is now 30. Although there are territories and pairs waiting to breed all around the island, those that have made a start are in clusters!
Diane Smith produced this map.
A couple of nests have had one or both eggs predated. The most intriguing incident of all left behind evidence that looked like this …
The nest had a single egg on several previous visits. The broken egg is still inside the nest cup. The obvious suspects are Mole Snakes, Kelp Gulls and Feral Cats. But Mole Snakes consume eggs whole; Kelp Gulls carry them away in their beaks; the mammalogists say this is not cat damage, and they suspect a stab by a beak. It turns out that there was another oystercatcher nest 5 m away. Bruno Ens, in The Netherlands, says that he has seen Eurasian Oystercatchers stab the eggs of their neighbours, so vandalism, rather than food, is probably the most likely explanation. The contents of the egg have dried out, rather than have been consumed.
The eggs in several nests have hatched since our last visit …
Newly hatched chick, still drying off. The eggshell, in two parts, has not yet been removed. Curated in observation.org
One of the views from the 360 degree office. Spring low tide. The black/dark brown stuff on the shore and in the sea is the kelp Ecklonia maxima, sea bamboo.
Bird list
This is a list of species recorded during this fieldwork trip. They are listed in the order recorded, with a list that started at the research house. Species in red are linked to the structured texts on the BDI website. Species with an asterisk have photos; they are below the table, and in the same order as listed.
* c. 300 about 2/3rds of way back to The Waterfront
Kelp Gull incubating.
Two feathered Kelp Gull chicks. The chick on the right still has a downy head.
Juvenile Eurasian Whimbrel. The pattern of feathers on the back gives this away. This bird is likely to be about six months old. Likely to have hatched in the taiga in Russia, the huge belt of coniferous forests between the tundra and the steppes.
Pied Crows.
Chukar Partridge, one of the parents of the chicks below.
These fluffballs could not be more than a day or two old. But they were FAST. This record is in observation.org as record 3817981025
Ring-necked Dove. Confirmed by the white tail in flight.
Crowned Lapwing chicks, on the football pitch at the maximum security prison, photographed through the two prison fences!
Sandwich Tern. It already had five or six new primaries on its right wing, and the 10th primary has broken tips and is bedraggled through a year’s wear. But the bill tip has very little yellow, and there are darker grey feathers on he wing coverts, so maybe it is a second year bird, i.e. hatched about May-June 2024. Ring recoveries show that most of the Sandwich Terns that migrate to southern Africa breed along the coasts of the North Sea.
Sabine’s Gulls seem to have become increasingly infrequent in Table Bay in the past two decades. We passed a substantial-sized flock. Geolocator studies show that the Sabine’s Gulls that migrate here breed in eastern Canada. There is a different set of photos in observation.org
Reptiles
There are lots of Angulate Tortoises on Robben Island. Mostly they are invisible. Then there is a cool breeze, or mist blows in from the ocean, or there is dew on the ground in the early morning, or there has been rain, and vast numbers are on the move. They risk their lives crossing roads. This one moved at speed.
This was the only Mole Snake on this trip. It is in observation.org
Fieldwork 3 : 27 and 28 November 2025
We found 14 new nests (S07 to S12, and N06 to N13), so the total number of nests found with eggs for the breeding season so far is 25. Nest S05, which had one egg on the previous trip, now has two! All nests found with eggs on previous trips still have the same number of eggs (or more!), so there have been no losses due to predation, or any other cause. In addition there is one pair which has two chicks, and a second pair suspected to have at least one chick.
Our study species, the African Oystercatcher. The bird on the left has been ringed.
Diane is measuring and weighing the eggs at nest S08. The process takes a few minutes, and is only done once. We have never had a desertion.
Nest S09, with two eggs, is in the centre of this photograph. There is also an arrow on the flat rock which points to the eggs.
Team 3 at work in the office.
Photogenic species!
The final stage of production of an oystercatcher egg is done in the paintshop. Every egg has a unique pattern.
This is the distribution of nests around Robben Island, 28 November 2025! The gap is the harbour. There appears to be strong clustering of nests.
Bird species27–28 November
This is our list of bird species, in the order in which they were recorded for the bird atlas project (SABAP2, pentad 3345_1820, Robben Island). The first bird seen after we started atlasing was a Chukar Partridge. There are some surprising omissions. We didn’t see a Cape Wagtail, although, between the four of us, we covered the entire coastline. We failed to see sparrows, neither Cape nor House. Pied Crow is missing. We looked for White-throated Swallow as we waited about 20 minutes at Murrays Bay Harbour for the ferry, but didn’t see them. The Bank Cormorant is the final species on the list; there were nests of breeding birds on the short arm breakwater as we left the harbour.
African Penguin was low down on the list at 17, i.e. the 17th species to be recorded. That is scary low. Most of these are juveniles doing their first moult. They moult into adult plumage, even though it is likely to be several years before they start to breed.
Cape Cormorant with chick. The number of Cape Cormorants breeding this spring seems much smaller than in previous years.
White-fronted Plover.
Other species
This big male Cape Fur Seal was one of three seen on the shore a couple of hundred metres south of the Faure Jetty. It has a nasty open wound on its right flipper.
We found this piece of shed mole snake skin at these coordinates along West Perimeter Road. The snake had clearly used a big piece of chipboard as den.
Cricket.
Sea urchin.
Abalone shell.
A lighter note, but actually dark!
73 lighters found on 2 km of shoreline by three observers! Robben Island sits alongside busy shipping lanes into the port of Cape Town, and also in the dispersal fan of the trash that washes out to sea from 214 square kilometres of the city of Cape Town via the Black River. See Industrial biodiversity 2 : Black River in Paarden Island : stormwater drain.
Fieldwork 2 : 18 and 19 November 2025
This was the second trip of the breeding season. Rene Navarro did the monitoring single-handed. He found nine new nests (S03 to S06 and N01 to N05). The two eggs in N01 are both pipped, so they were laid about a month ago! S01 and S02 (see Fieldwork 1, below) are still going strong.
S03. First nest of the Fieldwork 2 trip. Minimalist approach to nest architecture.
Both eggs in nest N01 were starred. The chicks will gradually turn the fine cracks into a bigger and bigger hole in the eggs, and will hatch within a couple of days.
Fieldwork 1 : 10 and 11 November 2025
We started the African Oystercatcher monitoring on Robben Island on 10 and 11 November 2025. The focus of this phase of the monitoring is on the 2025/26 breeding season.
Theoretically, it is exceptionally early in the breeding season. The African Oystercatchers should be right at the start of their breeding season. To our astonishment we found that one pair were the proud and noisy parents to two recently-hatched fluffballs:
The chick at the bottom of the photo is not yet completely dry, so had hatched in the previous couple of hours. The nest, i.e. the place where the eggs were incubated. is the shallow depression immediately to the left of this fluffball.We knew there were chicks because the parents were going absolutely ballistic. They are totally blasé when there are eggs. But the moment they hatch, there is a radical change in behaviour.
One of parents pretended to be injured. It did an Oscar-winning performance, a distraction display in which the “injured” bird tries to lead you away from its chicks. As in the video below!
The video failed to capture the more determined parts of the display.
One of the parents was ringed:
The ring has six characters. The left photo shows: K50 _ _ _ ; the right photo gives _ _ _ 313. Therefore K50313. It had been ringed, as a breeding adult, on 4 January 2019. Potentially, we can read a more ring numbers by taking lots of photographs. Say, 30. Then you can hope to get an in-focus photo of each digit!
We saw lots and lots of scrapes, some of which will have eggs added over the next weeks and months. We found two nests with eggs: S01 and S02. Both had two eggs. Here are photos of the nests.
S01 is a scrape that has been neatly line with shell fragments.
S01, the “Traffic Beacon Nest”. The big marker is a few metres away from the nest. Close to the nest there are arrows on the rocks to point to the nest. If the nest gets destroyed, by a predator, or worse, a storm (which moves stuff around), we can then work out where the eggs actually were. We need to do this, because the African Oystercatcher takes a minimalist approach to nest architecture.
S01 is a nicely concealed nest, above the spring high tide level.
S02 is a little depression, lined with fragments of shells and little stones, just on the seaside of a small ridge of kelp. The incubating oystercatcher can see over it, but its body is concealed.
The “Box Nest” has a series of markers, starting at the road around the perimeter of the island.They get more subtle closer to the nest.
The final marker is the little piece of white plastic (there is lots of debris washing up on the shore) halfway between the box and the nest, just at the thin line of kelp.
Mammals: Cape Golden Mole (fresh runs), Humpback Whale (three together, including a small calf, just outside Murrays Bay Harbour on 11 November), Steenbok.
Butterfly: Cabbage White
A Sunfish was seen during the trip back to the Waterfront.
Papers
We have done a fair amount of science over the years resulting from this monitoring. Here are some of the Open Access papers.
Braby J, Underhill LG 2007. Was poor breeding productivity of African Black Oystercatchers on Robben Island in 2004/05 caused by Feral Cats, Kelp Gulls, Mole Snakes or the Sumatra tsunami? Wader Study Group Bulletin 113: 66–70.
Calf KM, Underhill LG 2002. Productivity of African Black Oystercatchers Haematopus moquini on Robben Island in the 2001/02 breeding season. Wader Study Group Bulletin 99: 45–49.
Quintana I, Button R, Underhill LG 2021. African Oystercatchers on Robben Island, South Africa: The 2019/2020 breeding season in its two decadal context. Wader Study 128: 209–219.
There are papers to be added. It will take a while to find the online versions.
Acknowledgements
We are grateful to the Robben Island Museum for a vast amount of logistical support for the African Oystercatcher monitoring project. This has been provided over a period of 25 years; we started in the summer of 2001/02. This fieldwork is done with ethical clearance and a research permit.
“How do we get people interested in the environment?” “How do people get connected to nature?” This gallery of photos, taken at the Nuwejaars Wetlands this past week, has one of the answers!
Beware. There is a mist net ahead. One of the poles is an arm’s length to the right of Rocco’s right arm! It took them by surprise.
Dembo, from The Gambia, is explaining Malachite Sunbird to Grade 7s. This was the oldest class at Elim Primary School. We had Grade 1 to Grade 7, plus the special needs school, split into 27 groups of about 10 in a group.
This male Malachite Sunbird has impressive pectoral tufts.
Diane explaining Cape Sparrow to a focused group of learners.
In training as a releaser of Fiscal Flycatcher. Note the correct ringer’s way to hold a bird.
There can be few activities that achieve “connectedness to nature” as well as bird ringing does.
We are grateful to the Nuwejaars Wetlands SMA (Special Management Area) for the opportunity to help with their environmental education programme.
This modest building is the nerve centre of all the activities of the Nuwejaars Wetlands SMA.
This was our first Cape Sugarbird at Hazevlakte. Our normal stake out for sugarbirds is about 8 km away. In contrast, Malachite Kingfishers are common in the floodplain.
Mist nets in the floodplain.
African Rail, a bird species that is not often seen! We handle them quite regularly here!
This was a week when mist netting was not easy. Spike Milligan has a poem called Granny. The poem has three verses, and the second verse goes like this.
All through the night the wind grew worse
It nearly made the vicar curse
The top had fallen of the steeple
Just missing him (and other people)
Our version of it runs like this:
All through the night the wind grew worse
It nearly made the ringers curse
The nets were moving as fast as arrows
Just scaring birds, like all the sparrows
So we spent some time looking at other things …
Raucous Toads Sclerophrys capensis making a new generation of Raucous Toads.
If you get the right angle on the hairy caterpillar of the Cape Lappet Moth Eutricha capensis it is delightfully attractive!
We found two damselflies. Common Threadtail Elattoneura glaca (top) and Common Citril Ceriagrion glabrum (bottom).
And we spent some time eating.
That looks like a good snack for lunch!
The pizza oven did good service again …
Roasted for an hour.
We had lots of time to talk, the Hazevlakte units have lots of comfortable space to do this. We planned a couple of papers for Biodiversity Observtions.
You cannot see the wind in a photograph!
For the first two mornings the wind was calm. The sheep did lawnmower duty, and the eucalypts reflected in the Nuwejaars River.
As we left the Agulhas Plain, we saw this fire starting in the outskirts of Bredasdorp.
All through the day the wind grew worse
It really made the fireteams curse
The flames were eating up the trees
and racing through the veld with ease
The fire was still burning along the top of the range of mountains west of Bredasdorp two days later! The choppers try to prevent loss of property.
Thanks, Ross, for the video.
There is a list of all the upcoming BDI events here. Links to reports of previous events are here. There is a blog about the value of bird ringing to research and conservation here. This blog is about the value of bird ringing as a way to develop connectedness to nature.
There is no pollution from city light. We are deep in the countryside. There is no moon. The night is dark. Above our heads, the Southern Cross and The Pointers remind us that we are in the southern hemisphere. To be precise, we are at Ouberg Private Nature Reserve, deep in the mountains behind Montagu …
Too soon, the night gives way to dawn, the ringers are up early, …
… checking the mist nets which were opened at first light …
… and which are essentially invisible against the background of vegetation.
This Acacia Pied Barbet is being held with its head between the first and second fingers, and then the back fits snugly into the palm of the hand. That beak is powerful and sharp! If you don’t hold a barbet correctly in the ringers’ grip, the head swivels round on the flexible neck, and the ringer’s blood starts to flow.
With an average mass of 6 g, the Fairy Flycatcher is the lightest (and smallest) bird species in southern Africa.
It is ringers who are privileged to see the delicate shade of peach on the underparts.
Pearl-breasted Swallow.
This is a list of species handled, and the numbers of each species. Those in red have links to structured species descriptions on the BDI website:
To what extent do birds in a habitat like this make use of the drainage lines as highways? That is a question that interests us!
This is the communal area at the campsite (glamping and bring-your-own-tent) at the Ouberg Private Nature Reserve. It is ideal for our needs.
One corner of the communal area at Ouberg Private Nature Reserve turned into ringing office!
These ringing blogs often lapse into a focus on food. This one is no exception. Two interns from the Basque Country in Spain, Ana and Iñaki, treated us to tortilla de patatas, aka, Spanish omelette.
You can get links to all our reports on events like this on this website. There is a list of future events here.
There is a description of the value of bird ringing to research and conservation here.
The Ouberg Private Nature Reserve is 20 km northeast of the town of Montagu in the Western Cape. It is in the foothills of big mountains. Three biomes meet here. The vegetation is a mosaic of Fynbos, Albany Thicket and Succulent Karoo.
Part 1 : The birds and the ringing
This is one of the Biodiversity and Development Institute’s core ringing sites. We have been ringing here several times a year since 2020. We are steadily building up a population of ringed birds, and we plan to analyse the data we collect. For example, there are no estimates of survival rates for most of the species that we ring. That needs a long-term ringing commitment to a site. There is a blog on the value of bird ringing here.
We used mainly mist nets, and we also had spring traps.
We handled a lot of interesting birds!
This Bar-throated Apalis hatched this past spring breeding season, so it is probably three or four months old. It is a young bird because it does not yet have a well-developed throat-band, and a dark-grey iris, rather than the white of the adult. The eye colour is not mentioned in the field guides!
This Acacia Pied Barbet is being held in the ringers’ grip, with the neck between the first finger and the second finger. In this position the bird fits gently against the front of the hand but under complete control. There is no chance of being bitten by that formidable beak, which can draw blood if the bird is not held properly. These are skills you learn as a ringer.
The Sombre Greenbul is not an easy bird to see, because it skulks in the canopy. So when it is in the hand, we get the opportunity to see the creamy white eye. The eyes of a bird appear to be on opposite sides of the head, which gives them a vast field of view. Imagine a circle of vision for each eye. From the photo above, we can tell that these two circles overlap a bit. When the bird looks at an object in this overlap zone, it can see properly in three dimensions, like we do. That makes it possible to guage the distance to fly to a branch, or to the next item of food.
It is only ringers who really get to see the royal purple patch on the rump of a White-backed Mousebird.
Cape White-eye. The white ring around the eye consists of feathers. Whereas for the …
… Cape Bulbul, the white around the eye is bare skin. This bare skin forms an ellipse, and the eye of the Cape Bulbul is a bit to the rear of the centre of the ellipse. In contrast, the African Red-eyed Bulbul has an eye-ring which is circular (and, obviously, red) and the eye is in the centre of the circle.
This Bokmakierie was a retrap! 4B16627 was ringed on 23 April 2025, during our last visit here.
And so was the next Bokmakierie! 4A93536 had been ringed here more than four years ago, on 24 July 2021. It has not been retrapped on any of the about 10 ringing visits we have made to Ouberg since July 2021.
Have you ever wondered how the white V on the back of a Southern Fiscal is constructed?
The table below shows that we handled a total of 138 birds, of which an impressive 28 were retraps. The retraps are especially valuable for the statistical analyses which estimate survival rates. But we need to keep this effort up for a bunch more years! Species in red have links to structured species accounts on the BDI website.
There is a list of future planned ringing events here. And a list of all past events which do have blog reports is here.
Part 2 : The food
Let the pictures tell the story!
Part 3 : The place
In 2022, Sue Gie made a presentation about the Ouberg Private Nature Reserve at the annual conference of the Southern African Wildlife Management Association (SAWMA). You can listen to it here (15 minutes). It provides a fascinating backdrop to help you understand the history and conservation value of Ouberg.
The photos below show the campsite, one of the four glamping tents, and the open-plan kitchen-dining-living space.
Ouberg Private Nature Reserve has a website. It represents an awesome place at which to escape from the city. The email address is info@ouberg.co.za.
The BDI is holding an Open Day for bird ringing at Grotto Bay on Saturday, 29 November 2025 15 November 2025. We will be hosted by the local community at Grotto Bay Private Nature Reserve. The reserve has an area of 385 ha, and the vegetation consists of Strandveld and Sandveld. It is about 60 km north of Cape Town, turning off the R27. We meet at the Conservation Centre within the reserve.
Two of our previous bird ringing visits to Grotto Bay have blogs describing the events: they were on 27 May 2023 and 31 August 2024. The privilege of being a ringer is the opportunity to see birds like this Long-billed Crombec up close, and to see the detailed structure of the feathers. There is a description of the value of bird ringing here.
Long-billed Crombec, ringed at the Open Day for bird ringing at Grotto Bay Private Nature Reserveon 31 August 2024.The Long-billed Crombec has a remarkably short tail. It is only when you see this bird in the hand that you can actually see the size of the tail feathers. Photo : Suzette Vorster
There is lots of space, so ringers are welcome to bring their own nets and ringing equipment. Afterwards we will make a total list of all birds ringed, and produce a blog for the event.
The reserve is not open to the public, so you will not just be able to arrive on the morning of the event! You will gently be turned away with 100% certainty! If you plan to attend, please send an email to me at les@thebdi.org before Tuesday 25 November 11 November. In the email, please include the registration of the vehicle you will travel in, plus ID number and cell phone number. That information is used to pre-arrange entrance at the gate. Information and a code of conduct for visitors to the Grotto Bay Private Nature Reserve is here.
Sunrise on 29 November 15 November is at 05h28 05h35. So the best time to arrive if you are planning to put up mist nets is around 05h00. Otherwise come anytime early in the morning; 07h30 to 08h00 is probably about optimal! We will probably have left by 12h00.
One of the most important components of being trainied as a bird ringer is to learn the skill of being able to extract birds from mistnets safely. This Cape Robin-chat was ringed on 31 August 2024. We might well retrap this bird on 15 December 2025. Photo : Suzette Vorster
There is a list of all the upcoming BDI events here. After Grotto Bay, the next Open Day for bird ringing will be on 20 December 2025, at Zandvlei. This will be ringing followed by brunch.
Links to reports of previous events are here, and there is a blog about the value of bird ringing to research and conservation here.
This was the first bird ringing visit to the Nuwejaars Wetlands SMA (Special Management Area). Our objective was to start the process of finding good ringing sites. We had several failures and a few successes!
One of the biggest successes was the discovery of the accommodation on the farm Hazevlakte. This is the view of the Nuwejaars River looking upstream …
… and below is the view looking downstream …
The eucalypts on the opposite bank are alien. Their redeeming feature is that they are used by African Fish-eagles, cormorants, etc.
The werf at Hazevlakte provides a variety of habitats for mist netting. We found that the species diversity extends beyond weavers, bishops, starlings and sparrows to include Blacksmith Lapwing. Southern Fiscal, Fiscal Flycatcher, Olive Thrush, Malachite Sundird and Cape Robin-chat.
Here’s a … thanks, Ryan
mist net being set up in a patch of grass on the tree-lined edge of the werf at Hazevlakte on a misty morning. There are little grove of trees to the left of the photograph. Weavers, starlings and sparrows were nesting in these trees, and flying out to the irrigated fields in the background to feed.
More mist nets being set up in the werf. Weaver nests in the background.
Male Malachite Sunbird, displaying one of his bright yellow pectoral patches.
Southern Fiscal, a male because of the absence of a brown patch on his flanks/
This Lesser Honeyguide was not in primary moult. Not surprising, because moult starts after the spring breeding season. Lesser Honeyguide is one of a tiny number of South African species with modelled estimates of the timing of moult, based on data collected during bird ringing. The mean starting date is estimated to be 8 January, and the duration is 140 days, so that 28 May is the mean completion date. There is a lot of individual variation. 140 days to replace primaries! That is four-and-a-half months. That is a pretty slow growth rate.
So the werf at Nuwejaars was declared a Potentially Important Ringing Site.
The werf at Hazevlakte has this negative bird that makes you wonder if your eyesight is going wonky. Unfortunately, we didn’t catch it, or any other Hadeda! [**Update : this leucistic Hadeda Ibis has been seen on almost all of our subsequent visit to Hazevlakte, and most recently on 22 September 2025**]
We did an exploratory trip one afternoon to a protea patch on the farm Zandvlakte. We caught four Cape Sugarbirds. These are quite likely the southernmost sugarbirds ever to be ringed! This locality was added to the list of discovered sites to be revisited on future trips, a second Potentially Important Ringing Site.
We tried several other sites, but none of them reached the threshold to become Potentially Important Ringing Sites.
There were days when we caught the sunrise, and little else!
The table gives the total number of birds of each species handled.
One of the six Malachite Sunbirds we ringed was a nestling at one of the less profitable sites! The date was 11 October 2024. This bird was found in a garden of a house near the southern tip of Africa, in L’Agulhas, 22 km from where it had been ringed. It was caught by a domestic cat, but not killed outright, but did not survive. Even though the story is tragic, it is an interesting example of juvenile dispersal.
Damselfly discovery
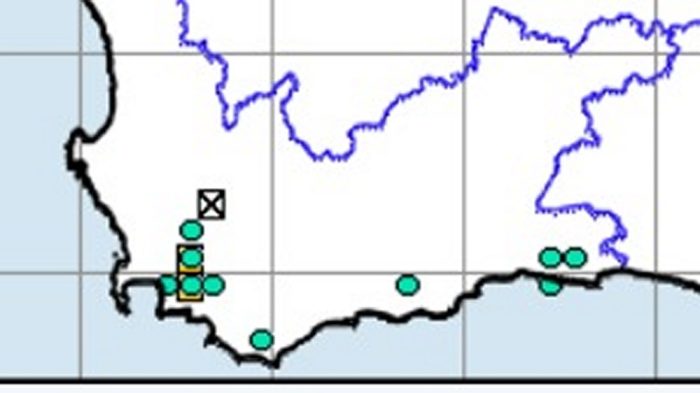
We did a ringing expedition to a site which turned out to be the poorest a site could possibly be. We had zero birds to ring. But we did have a damselfly to photograph. To everyone’s astonishment, the damselfly was identified as Spesbona. This species is classified as Endangered; it is extremely rare and localized. Before looking at the damselfly, look at the distribution map for the species …
The new record is the southernmost, and a long way from the others, which are along the mountain chain between Knysna and Somerset West.
This is the Spesbona seen at Hazevlakte on the Nuwejaars Wetlands. In relation to most other species of dragonfly and damselfly, this is a species that is in flight relatively early, with a mid-October peak. You can read up about Spesbona in the Online Atlas of Dragonflies and Damselflies of South Africa, with the species text here.
Thanks
Erica Brink, Nuwejaars Wetlands Special Management Area, organized our accommodation with Karen Neethling at Hazevlakte. It turned out to be an inspired arrangement. Eugene Hahndiek helped us find ringing sites, and arranged access to them.
We are reporting on our eighth bird ringing expedition to the Nuwejaars Wetlands. We were here from Friday 19 to Tuesday 23 September 2025, four nights. As with the previous visits here we are still exploring new sites and new options. Some are winners, and some are less successful. But it is all part of the learning process.
Saturday morning was spent at the northern side of the Nuwejaars River floodplain at Moddervlei. We didn’t get the mistnets in good places and a stiff breeze started early. We could have done better here. On Sunday morning we put the mistnets up near the hide on the African Marsh Harrier Trail. This is now our best site in the area! On Monday morning we ringed around the werf at Hazevlakte, around our accommodation, and that yielded a steady stream of birds, lots of them retraps. On Monday afternoon we reccied a spot with easy access to the Nuwejaars River close to the farm Heuningrug. Definitely worth a bigger exploration. On Tuesday morning we operated near the protea patch at Zandvlakte. This is a regular spot, and the place where we hope to get samples of sunbirds and sugarbirds.
As the days pass by around the spring equinox, the rate of change of sunrise is fastest. and in the wrong direction! So, for the next couple of months, the ringing day starts a minute of two earlier every day.
06h02 in the Zandvlakte protea patch. Time to get the mist nets up, watched by the “morning star”, in this case, Venus.
Central to every bird ringing day is the table where the ringing gets done. It’s chosen to be out of the sun if possible (otherwise we need to take the lapa for shade). It mustn’t be too far from the mist nets, and it must not be too close either. The global bird ringing standard is to keep birds in bags from capture to processing. And we keep the bags in order so it is a FIFO system. FIFO = First In, First Out. Most of the time the designated ringer doesn’t know what is in the next bag until he takes the bird out.
Bag 1
What’s in the bag? It’s a …
… Cape Sugarbird. The first of seven that we ringed at Zandvlakte, in the protea patch. That brings the total number of sugarbirds ringed at Zandvlakte to 14.
Part of the Zandvlakte protea patch. These are cultivated proteas, planted in rows, but clearly many (in reality, most) of the plants didn’t survive.
Even though it is a cultivar, a horticultural variety of pincushion protea …
… it still produces nectar for the sugarbirds.
The sugarbirds make excursions into the cultivated patch of proteas from the neighbouring hillside, which is covered by a remnant of natural fynbos.
Bag 2
Here is another bag with a bird in it …
This bag comes from the recce trip to the Nuwejaars River on Monday afternoon …
… and the bird in the bag is a Black Crake, our third for the trip! The floodplain of the Nuwejaars River has a large population …
… and here, just before release here is the crake with its habitat in the background. To see the colours of the eyes, bill and legs with the bird-in-the-hand is an awesome privilege.
More privileges …
This is the southernmost Diederik Cuckoo ever ringed …
It’s a male, diagnosed by the red eye-ring
It is only ringers who really get to see birds close-up!
Here is the eye of the Red-eyed Dove.
This is the tail pattern, which you can compare with the tail pattern of the Ring-necked Dove ….
… in which the outer three tail feathers are white, but which can be hidden away when the tail is in the natural position.
… this is a young Fork-tailed Drongo. The white pattern of the underparts, and the dark eye (red in adults) are diagnostic of this being a juvenile.
Summary table
Dieter Oschadleus produced this summary of the bird ringing at Nuwejaars Wetlands in September 2025.
It is still a bit early in spring for dragonflies. This is the commonest species in the Agulhas Plain, the Red-veined Darter. This is often one of the first species to be on the wing in summer. Its range covers almost all of Africa, southern Europe, and on into Asia.
The Painted Lady has the largest distribution of any species of butterfly. Here is the evidence that it extends to the southern tip of Africa. It some parts of the world it is a migrant, for example from Africa to Europe.
Thank you
We are grateful for the support of the team at the Nuwejaars Wetlands Special Management Area, Ross Kettles and Erica Brink. Thanks to Karen and Con Neethling for the warm welcome, once again, to the accommodation on the farm Hazevlakte. Thanks to Con, Liohan Giliomee, Dirkie Swart and Diko Swart for permission to ring on their farms.
Annual report: Nuwejaars Wetlands Special Management Area
To understand what we are involed with, you can study the 2024/25 annual report of the Nuwejaars Wetlands SMA is available. Read it here.
… and finally …
… even if we were at the Nuwejaars Wetlands for with the sole purpose of being outdoors, it would be valuable therapy for mind and body!
A paper on the migration of Barn Swallows has been published Open Access in the journal Birds. Publication date was 9 September 2025. The authors, title and journal reference are
Underhill LG,Remisiewicz M 2025. Arrival and peak abundance of Barn Swallows Hirundo rustica in three regions of South Africa in relation to climate indices, deduced from bird atlas data. Birds 6: 48. https://doi.org/10.3390/birds6030048
You can find the paper here or if you would like to go directly to the pdf version, it is here.
In the paper we discuss the way in which the arrival and peak abundance of Barn Swallows in South Africa are influenced by large-scale climate indices in the areas in which they breed, the areas in which they prepare for migration and in areas along the migration flyways.
We, in this case, are Magda Remisiewicz and myself. Magda was a post-doctoral research fellow in UCT’s Animal Demography Unit from 2008 to 2010. She now heads the Bird Migration Research Station at the University of Gdańsk in Poland and has recently been appointed professor at this university. Part of the formalities in being appointed a professor at a Polish university is to be congratulated by the national president:
President Andrzej Duda congratulates Professor Magda Remisiewicz on her appointment as Professor, 24 June 2025
The data analysis for the paper uses Barn Swallow records on SABAP2 checklists collected at the three main centres where the most atlas data has been collected: the area around Cape Town, called Greater Cape Town in the paper; the area around Johannesburg and Pretoria, Greater Gauteng; the area around Durban, Greater Durban. For each of these regions there are lots of checklists for every one of the 17 years between 2007, when SABAP2 started, and 2023; we stopped then because we did the analyses for the paper in 2024. In this blog, we focus on the results for Greater Durban.
Calculating how early or late the Barn Swallows are each year …
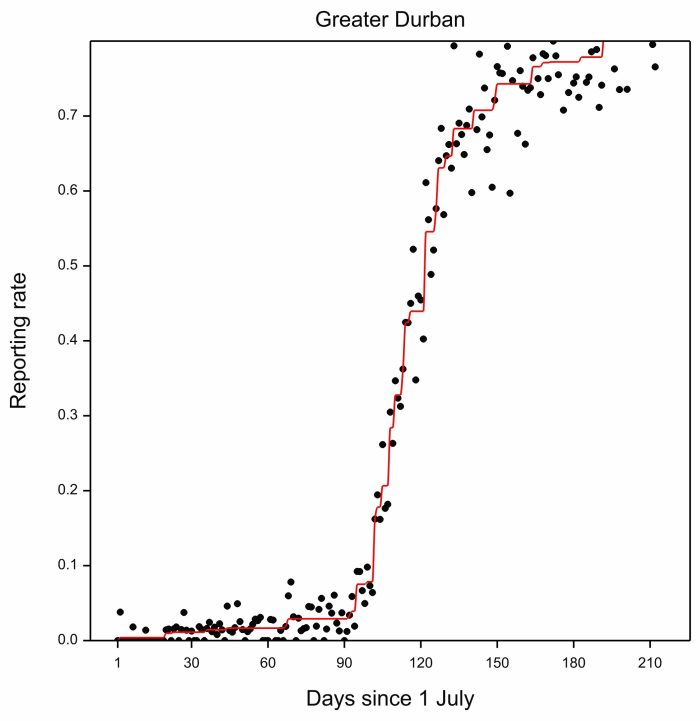
We started the analysis by combining the data for all 17 years for each region, and produced a curve which showed how reporting rates increased on average during the arrival periods in each region. Here is the plot for Greater Durban:
The black dots are the daily reporting rates in the Greater Durban area from 1 July. So the dot for 1 July would use the SABAP2 checklists made on 1 July in all 17 years (2007 to 2023). It is clear that the period of the most rapid increase in reporting rates runs from about Day 100 (i.e. 100 July) to around Day 130 (i.e. 130 July, known to most people as between 8 October and 7 November). The rate of increase of reporting rate then slows down, and flattens out by about Day 190 (i.e. 190 July = 6 January).
It is encouraging to see how well-behaved the pattern of daily reporting rates is. At the end of the day, that is a huge accolade to the quality of the atlasers.
The red line in the plot above is constructed using a technique called monotonic regression, which hugs the reporting rates as closely as possible, while always moving upwards. The red line defines the average timing of migration over the 17 years. For each year we want to find the anomaly of migration timing, ie quantify how early or how late migration was in relation to the long term average. And we want to do this over the whole arrival period. For Greater Durban we used the period between Day 104 and Day 192 as the arrival period (12 October to 8 January)
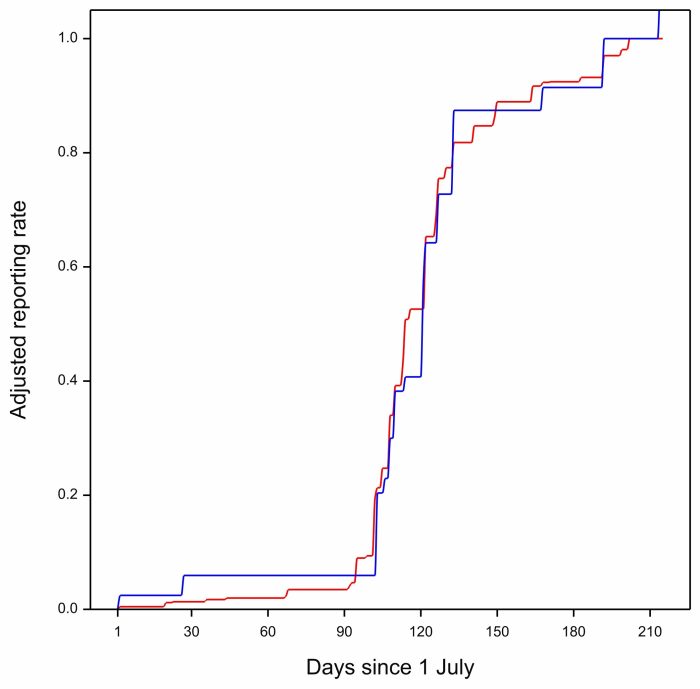
To calculate the anomaly for a particular year, we calculated the daily reporting rates for that year. Then we fitted the monotonic regression for the year. So for Greater Durban in 2016, the blue line is the line for the year, and the red line is the line for all the years, copied from the image above.
We adjusted the reporting rate curves for the individual years, (and for the multi-year curve) so that they ended at 1.0 in every year. This was done because the maximum reporting rate reached at the end of migration differed between years. In the curve above, for 2016, the red line (the multi-year curve), and the blue line (the 2016 curve) are very similar. The areas of the pieces of the blue curve which are above the red curve more or less balance with the ares of the blue curve below the red curve. In other words, arrival of Barn Swallows in 2016 had a very similar pattern to the overall arrival pattern, and the arrival anomaly for 2016 is small.
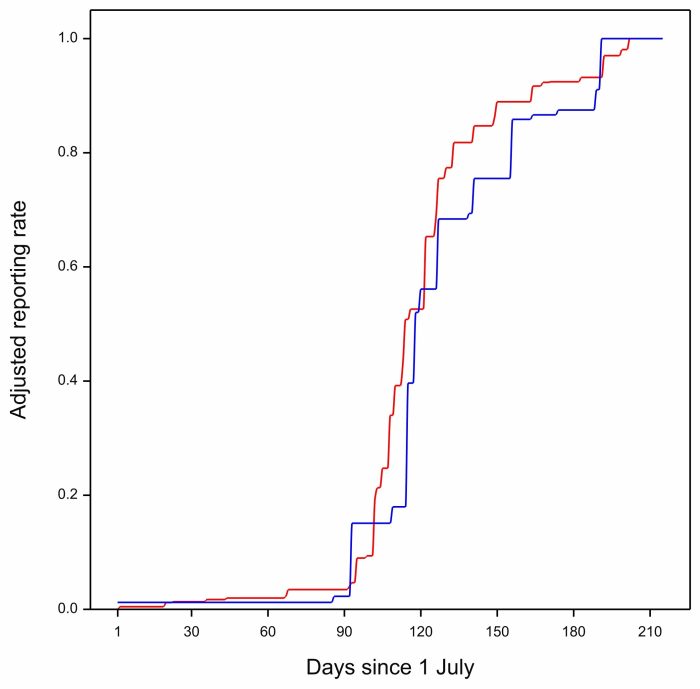
… next, have a look at this, the two curves for 2008:
The blue curve for 2008 is mostly to the right of the red multi-year curve. The Barn Swallows are late, and the anomaly is positive. In all of these plots the red curve is identical. It is the blue curve that represents arrival in a particular year and is different in each plot.
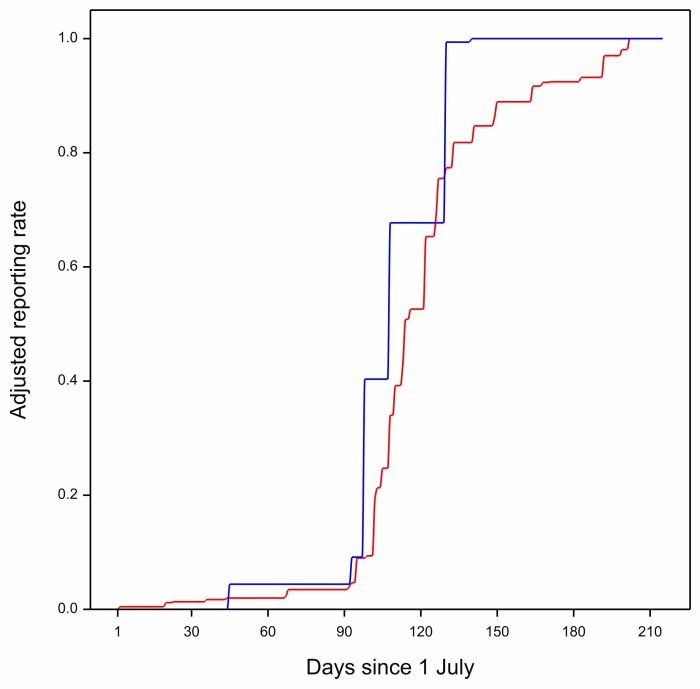
… and here is the curve for 2021:
The blue curve for 2021 is mostly to the left of the multiyear curve. Arrival is early. The anomaly for 2021 is negative.
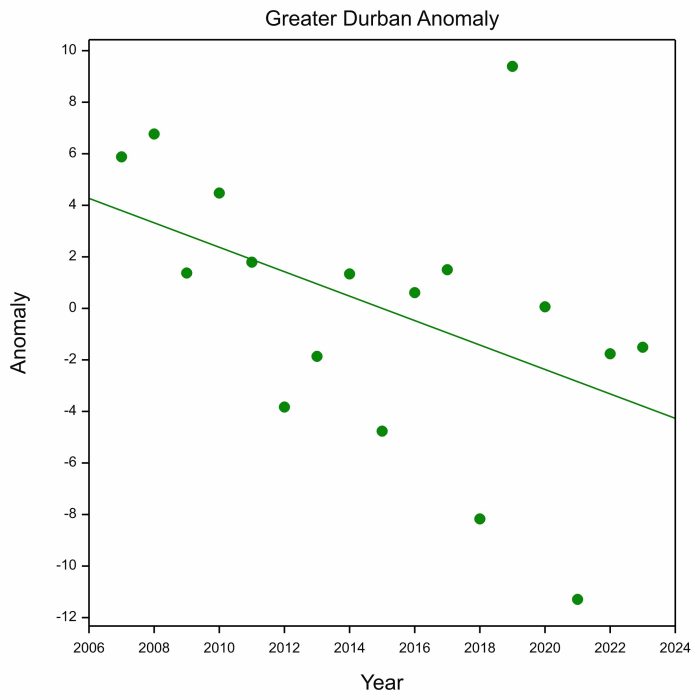
When you calculate the anomalies for Greater Durban for every year, and produce a plot with year on the x-axis and with annual anomaly on the y-axis, it looks like this:
The astonishing thing is that the scatter of points is not random, they point downwards. In the early years of SABAP2, the annual anomalies were mostly positive, and towards the end they were mostly negative. When the anomaly is positive, arrival is late, and when it is negative, it is early. So the downward trend in the plot above is saying that the arrival of Barn Swallows has trended earlier in KwaZulu-Natal during the SABAP2 period, 2007 to 2023.
The next result was even more surprising. These annual anomalies were positively correlated with a climate index called the Scandinavian Pattern. We used the average value of the Scandinavian Pattern during the breeding season of the Barn Swallows, the period May to August. Positive values for the Scandinavian Pattern are related to warm and dry conditions in northeastern Europe and western Asia, the breeding area of a large proportion of the Barn Swallows migrating to the Greater Durban region (we know this from ring recoveries). Favourable breeding conditions probably increase the breeding success of Barn Swallows, which can raise two broods. The adults with second broods, and the juveniles from them, will tend to migrate later than in years when conditions are cold and wet when the Scandinavian Pattern is negative. So the positive correlation between the timing of arrival on migration and the Scandinavian Pattern during the previous breeding season is eminently plausible.
Peak annual reporting rates …
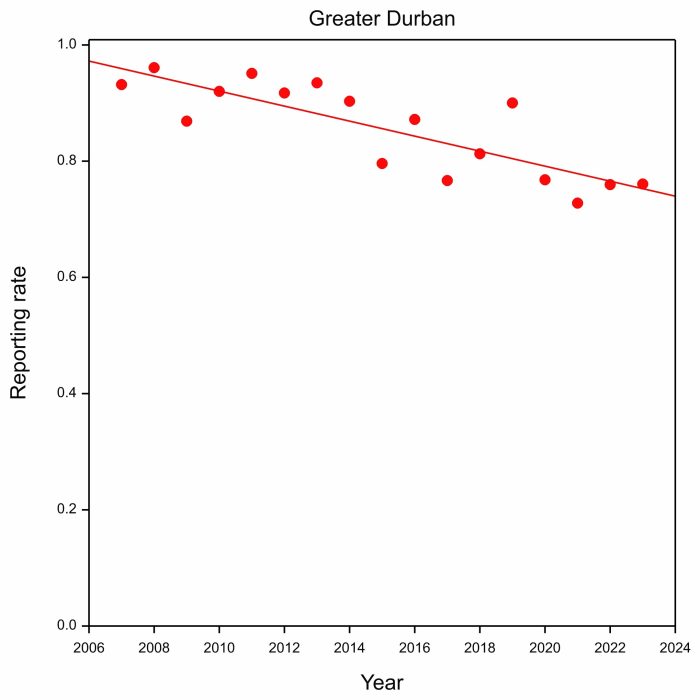
One of the things we discovered was that the reporting rates climbed to different values after each arrival year. So we computed the annual peak reporting rates for Barn Swallows for a given year as the median of the reporting rates during the midsummer period between 11 and 31 January of the following year. So the peak reporting rate for 2007 is based on the reporting rates for the three-week period between 11 and 31 January. We chose this period because reporting rates had stopped increasing by 11 January, and had not started decreasing by 31 January. So the plot below shows peak annual reporting rates for Greater Durban:
The results are alarming. They show a steady decrease in reporting rates from above 90% in the early years of SABAP2, to below 80% in recent years.
What are the take home messages?
The paper is packed full of results like this, relating to the arrival and peak abundance of Barn Swallows. Its main message is that the southward migration of Barn Swallows and the timing of their arrival at their southernmost non-breeding destinations are influenced by the big climate indices, and by temperature and precipitation across the breeding range and along the migration routes. The climate variables that show correlations make sense in relation to the known breeding areas and migration routes to the three destination regions.
The paper demonstrates the potential of the Second Southern African Bird Atlas Project (SABAP2) as a long-term annual monitoring project for birds. The particular strengths are its strong protocol, which enables the analysis undertaken here, and the fact that, unlike most monitoring projects, it enables monitoring to be done both in space and time. We recommend the indefinite continuation of the SABAP2 project, which with time, will provide more insight into changes in birds’ distribution and timing and their response to climate change not only in South Africa, but also on the breeding grounds and along migration routes. This might be the first paper to use SABAP2 data on an annual basis.
Barn Swallows arriving after sunset at an overnight roost in South Africa
Sister papers
This new paper has four “sister” publications, based on bird ringing done at the bird observatory at Bukowo, on the Baltic Sea coast in Poland. All of them are Open Access, Click on the journal reference to get the paper.
Remisiewicz M, Underhill LG 2020. Climatic variation in Africa and Europe has combined effects on timing of spring migration in a long-distance migrant Willow Warbler Phylloscopus trochilus. PeerJ 8: e8770. This paper has a blog.
Remisiewicz M, Underhill LG 2022a. Large-scale climatic patterns have stronger carry-over effects than local temperatures on spring phenology of long-distance passerine migrants between Europe and Africa. Animals 12: 1732.
Remisiewicz M, Underhill LG 2022b. Climate in Africa sequentially shapes within-season spring passage of Willow Warbler Phylloscopus trochilus through the Baltic Coast. PeerJ 10:e12964.
Remisiewicz M, Underhill LG 2025. Climate in Europe and Africa sequentially shapes spring passage of long-distance migrants at the Baltic coast in Europe. Diversity 17: 528.
These four papers deal with northward migration. The new paper deals with southward migration.
There is a full list of all publications with BDI authors here