We counted the African Oystercatchers on the 10 km of shoreline of Robben Island on 13 June 2026. There were 420 adults and three juveniles.
The previous survey of African Oystercatchers on Robben Island was less than a month ago, on 17 May 2026, and the total number counted was 479 birds. This is the kind of variability we experience between surveys. We don’t know whether the variation is due to our inefficiencies, or whether it is genuine, with a few tens of birds moving between the mainland and the island.
Besides the oystercatchers, we observed lots of other things …
… What are these guys focusing on? …
… two Humpback Whales
During past couple of months, the interior of the island has changed from brown to green. There has been excellent early winter rain in Greater Cape Town. There are already some flowers. The Kelp Gulls are settling into their colony round the northern edge of the island. But there seemed to be no nests as yet.
On the other hand, breeding for Hartlaub’s Gulls has been ongoing for several months through autumn. Not only are there nests with eggs, there are fledglings and …
… small chicks.
The Greater Crested Terns seem to near the end of breeding. Most of the juveniles that are remaining on the island can fly.
It is striking how much shorter the fledgling’s bill length is compared to the adult behind it. The chick in the front of the group can fly, but its bill has a lot of growing to do. This needs a consultation with Janine le Roux’s MSc thesis, on the growth of Greater Crested Tern chicks. Janine found that when a Greater Crested Tern chick hatches, the tarsus is already about 64% of adult size, and is almost completely grown halfway through the fledging period. Well-developed legs at hatching are essential, because the chick has to be instantly mobile. The wings start at 6% of adult size, and grow rapidly; the chick can fly when the wings reaches 72% of adult length. The bill lags behind in growth. It is 24% of adult length at hatching, and only 58% at fledging. This is a clever strategy, because once the chick can fly, it gets taken out to sea by its parents, and they feed it at the spot where the food is. No more flying long distance with fish-in-bill. This is an efficient system.
Other species seen during the count included this Ruddy Turnstone and this Grey Plover. It is June, and these species are about to start breeding on the Siberian tundra. The juveniles of these two species don’t breed until they are two years old. It is pointless, and risky, flying all the way to Siberia just to look around. So they spend their first “summer” in the southern hemisphere, and those of us who live in the southern hemisphere say they are “overwintering”, but a northern hemispherist would say they are “oversummering”.
This Northern Giant Petrel flew close inshore. This is an unusual sighting.
This Southern Fiscal carefully avoided perching on the razor wire at the prison.
Chukar crossing a road at speed!
The Sea Challenger went aground on the northern tip of Robben Island in May 1998, 28 years ago. It is one of the key landmarks on the perimeter of the island that we use for orientation. It was solidly built and is slowly starting to show the first signs of disintegration.
We are extremely grateful to the Robben Island Museum for their logistic support that enables this monitoring to be done.
On 24 May 2026, we were bird ringing at the Strandfontein Sewage Works at a different spot to where we were eight days ago, on 16 May! Relevant permissions were obtained beforehand.
These sewage works represents one of Cape Town’s most precious bird resources. The effluent flows from the main treatment operation through a large series of pans which attract a vast variety of birds. Especially during the dry summer months, the superabundance of water, generated by flushing loos and having showers, provides a critical drought refuge in a region where three centuries of development have destroyed or damaged many of the natural wetlands.
Conditions were near-perfect for mist netting. As the reflections in the photo above shows, there was little wind and it was overcast for most of the morning. We handled 80 birds of 11 species:
Three of the 80 birds we handled would qualify as waterbirds, on a strict assessment. These are the top two species on the list above. Seven of the remaining nine species would be considered as frequently associated with water, the exceptions being the Cape Robin-chat and the Southern Double-collared Sunbird.
Pride of place goes to the Black Crake, a reedbed skulker.
We ring Black Crakes above the “ankle”. The ringing pliers has slots of various size; the ringer chooses the appropriate size for the ring being fitted. The leg is held gently but firmly so that there is no chance of the leg being damaged as the ring is closed into a neat circular cylinder. The ring size is chosen so that it is loose, but not wobbly.
We also caught two Cape Teal …
… and that made up the three waterbirds, of two species.
There was an exceptionally late Brown-throated Martin breeding colony nearby, still active in May. In the winter rainfall area breeding peaks from September to December but has been recorded throughout the year.
This demonstrates the recipe for creating a breeding site for Brown-throated Martins. The near vertical sandbank is maintained by the thick tangle of vegetation at the top of the “cliff” which prevents the sand from slipping off, and a stream to keep eroding the base! The martins are exploiting the opportunity far beyond the limits of the “normal” breeding season.
An adult Brown-throated Martin in a mist net.
Newly fledged Brown-throated Martin
Another angle on the bird above, What gives it away as a young bird is pale-brown edging to so many of the feathers …
… including the feathers on the rump, the row of feathers forming the wing coverts, and the tips of the tail feathers.
Levaillant’s Cisticola with the Muizenberg Mountains in the background!
The Strandfontein Sewage Works is best known for the large array of waterbirds attracted there. Here is a tiny selection of common species …
… Little Egret in one of the shallow ponds …
… Glossy Ibis and Little Egret hunting for food together …
… Cattle Egrets on a bank between ponds …
.. Great White Pelicans flying over …
… and a Black-necked Heron on its way to the adjacent rubbish dump …
So that’s the wrap about our bird ringing at Strandfontein Sewage Works, 24 May 2026.
Cover image of Sombre Greenbul by Terry Terblanche – Addo Elephant National Park, Eastern Cape
Greenbuls belong to the Family PYCNONOTIDAE (Greenbuls, Bulbuls, Brownbuls, Leafloves, Bristlebills). They are mid-sized passerines with longish tails and mostly plain-coloured plumage. The family is distributed across most of Africa and the Indian Ocean islands, into the Middle East, through tropical Asia to Indonesia, and as far north as Japan. They occupy a variety of habitats, ranging from dense rainforest to arid savanna. The family comprises 161 species from 28 genera.
Identification
The cheerful Sombre greenbul has dull olive-green upperparts and olive-grey underparts with no other conspicuous features except pale whitish eyes. The Bill is black, and the legs and feet range from brownish to black. The sexes are alike.
Sombre GreenbulAndropadus importunus Gamtoos River Mouth, Eastern Cape Photo by Cobus Elstadt
Juveniles resemble adults, but have a paler bill and an orange gape. They also have brown eyes and an indistinct yellowish eye ring.
The Sombre Greenbul is easily mistaken for other greenbul species. However, the white eye, if seen, is diagnostic. The Stripe-cheeked Greenbul (Arizelocichla milanjensis) has a grey (not olive green) crown and face, and dark (not white to pale yellow) eyes. Yellow-streaked Greenbul (Phyllastrephus flavostriatus) has a grey (not olive green) crown and ear coverts, a whiteish (not olive-grey) throat, and brown (not white to pale yellow) eyes. The Yellow-bellied Greenbul (Chlorocichla flaviventris) has bright yellow (not olive-grey) underparts and red eyes.
Status and Distribution
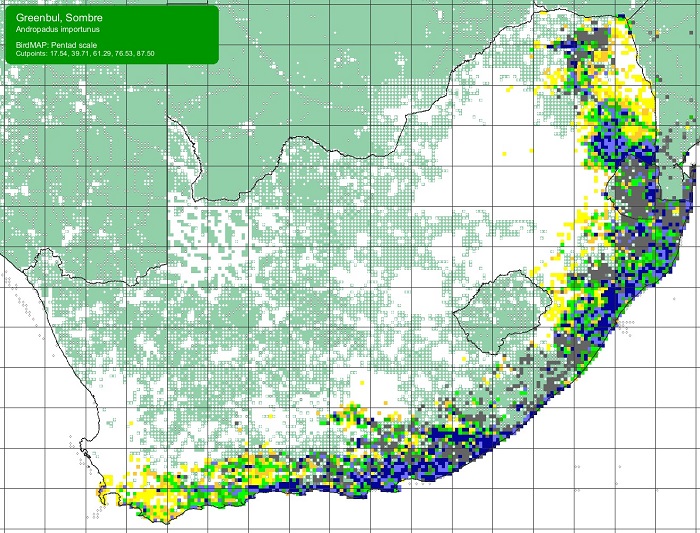
The Sombre Greenbul is common throughout its range. It ranges from Kenya and southern Somalia south to south-eastern Zambia, southern Malawi and South Africa. In southern Africa, the Sombre Greenbul occurs in eastern Zimbabwe and along the Zambezi River valley to Lake Kariba. It is also found throughout Mozambique, to northern and eastern South Africa, where it extends along the moist escarpment and lowlands south to the Cape Peninsula in the Western Cape.
SABAP2 distribution map for Sombre GreenbulAndropadus importunus – December 2024. Details for map interpretation can be found here.
The Sombre Greenbul is not threatened. Its range is not known to have undergone change since its discovery in the early 19th century.
Sombre GreenbulAndropadus importunus Cape St. Francis, Eastern Cape Photo by Gregg Darling
Habitat
The Sombre Greenbul inhabits Coastal scrub, coastal and afromontane forest, thickets in densely wooded valleys and various woodland types, including thornveld and dry woodland thickets along drainage lines. The Sombre Greenbul prefers natural vegetation, but is also found in alienthickets and in gardens in well-wooded suburbs. In the fynbos region, it is generally found around forest patches and fringing woody vegetation along rivers.
Typical Habitat Near Pietermaritzburg, KwaZulu-Natal Photo by Alex Briggs
Behaviour
The Sombre Greenbul is normally encountered singly or in pairs. Occasional large non-breeding flocks occur at abundant food sources. It is a resident and sedentary species.
Sombre GreenbulAndropadus importunus Maputo, Mozambique Photo by Pieter Cronje
It is shy and inconspicuous and is far more often heard than seen. Its ‘willy’ call is a well-known and characteristic sound wherever it occurs. It is best observed when loudly advertising its territory from the top of a tree or bush. Otherwise, the Sombre Greenbul keeps to dense cover, usually in the canopy. It is an inquisitive bird but dives quickly into cover when alarmed. The flight is rapid and direct, with frequent sharp swerves. It sometimes flies with exaggerated wing-beats, with soft wing clapping.
Sombre GreenbulAndropadus importunus Vleesbaai, Western Cape Photo by Pieter La Grange
The Sombre Greenbul spends most of its time in forest and thicket canopies, but occasionally feeds in the undergrowth, or on the ground among leaf litter. They sometimes also hawk flying insects, before diving back into cover to eat. The Sombre Greenbul consumes a wide range of wild and cultivated fruits and berries. It also eats insects, small snails, succulent leaves, flowers and unopened buds, and aloe nectar.
Sombre GreenbulAndropadus importunus Plettenberg Bay, Western Cape Photo by Itxaso Quintana
The Sombre Greenbul has a long breeding season in southern Africa, with egg-laying occurring in any month from October to April, but with a clear peak from October to December. It is a monogamous, solitary nester and strongly territorial. The nest is usually a flimsy open cup made from twigs, rootlets, grass, lichen and other plant material, lined with finer plant fibres and sometimes also hair. It is placed up to 4 m above the ground, in the main fork of a slender tree or shrub, or sometimes at the end of a branch and is usually attached to supporting twigs with spider web. The nest is built entirely by the female while the male sings from a nearby tree.
Sombre GreenbulAndropadus importunus Mkhuze Game Reserve, KwaZulu-Natal Photo by Lia Steen
1 to 3 eggs are laid per clutch at 1-day intervals. The incubation period lasts for 12 to 13 days, and is performed entirely by the female. The male brings food to her while she is incubating. Interestingly, the adults often perform a broken-wing distraction display when the nest is threatened, dropping directly to the ground and scuttling away like a rat. The nestling period lasts for a further 10 to 14 days, and both adults feed and brood the chicks. Both parents are recorded to eat the nestlings’ faecal sacs. The adults continue to feed the young for up to 3 weeks after the fledglings have left the nest. Broods of the Sombre Greenbul are frequently parasitised by the Jacobin Cuckoo (Clamator jacobinus).
Sombre GreenbulAndropadus importunus Vleesbaai, Western Cape Photo by Pieter La Grange
Further Resources
This species text is adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Alex Briggs, Cobus Elstadt, Dieter Oschadleus, Gregg Darling, Itxaso Quintana, Lia Steen, Pieter Cronje, Pieter La Grange, and Terry Terblanche is acknowledged.
Recommended citation format: Tippett RM. 2026. Sombre Greenbul Andropadus importunus. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/06/03/sombre-greenbul-andropadus-importunus/
List of bird species in this format is available here.
Cover image of Brown-throated Martin by Pamela Kleiman – Ukhahlamba Drakensberg Park, KwaZulu-Natal
The Brown-throated Martin belongs to the family HIRUNDINIDAE, a group that includes Swallows, Martins, and Saw-wings. They are small to medium-sized passerine birds, noted for their streamlined bodies, long, pointed wings, and square or forked tails, often with very long outer rectrices. Their bills are small and flattened with a wide gape, and their legs are very short and weak. All are highly adapted to aerial feeding, and most are gregarious, at least when not breeding. This family has a global distribution and comprises around 90 species, with the greatest diversity being found in Africa. 21 species have been recorded from southern Africa.
Identification
The Brown-throated Martin is a small swallow attaining a length of 12cm and a weight of 12.5 grams. The sexes are alike.
Brown-throated Martin Riparia paludicola Midmar Nature Reserve, KwaZulu-Natal Photo by Malcolm Robinson
It is found in 2 colour morphs. Adults of the more common pale morph are overall brown to greyish-brown, except for the belly and undertail coverts, which are white. The extent of white on the belly is variable. The uncommon brown morph is the same but has a brown belly and undertail coverts. At rest, the folded wing tips extend beyond the tail. Juveniles resemble the adults of their respective morph, but the feathers of the upper parts, especially wing coverts and rump, have buffy tips, and the underparts are washed rufous-buff.
The Brown-throated Martin is readily identifiable. However, the Sand Martin (Riparia riparia) is similar, but has a white throat and a brown breast band. The larger Banded Martin (Riparia cincta) has a distinct white supercilium and white (not brown) underwing coverts. The Rock Martin (Ptyonoprogne fuligula) is also larger, with contrastingly pale brown underwing coverts and white spots in the tail.
Brown-throated Martin Riparia paludicola Carnarvon district, Northern Cape Photo by Ryan Tippett
Status and Distribution
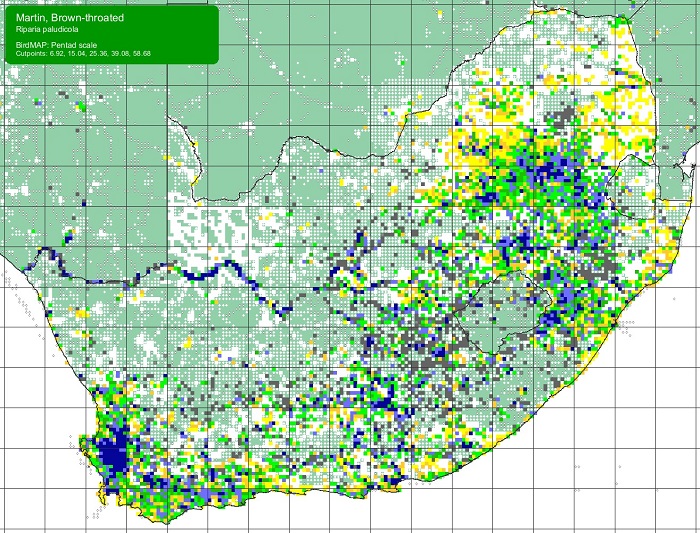
The Brown-throated Martin is locally common in southern Africa where it is widespread, especially in the moister northern, eastern and southern parts. It is sparse throughout Namibia (except the Caprivi Strip), and is largely absent from the Kalahari Basin. In arid areas, it is concentrated along large rivers, most notably the Orange River in the Northern Cape. This species has an extensive breeding range throughout Africa and Madagascar, through southern Asia and as far east as Taiwan and the Philippines.
SABAP2 distribution map for Brown-throated MartinRiparia paludicola – December 2024. Details for map interpretation can be found here.
The Brown-throated Martin is not threatened. It has probably benefitted from erosion, road cuttings, sandpits and drainage pipes that may have increased nest site availability. The widespread manipulation of natural wetlands by humans is likely to have had significant effects. Artificial impoundments, wetland drainage, and the regulation of river flow are all likely to have had substantial positive or negative impacts on local populations.
Brown-throated Martin Riparia paludicola Vanryhnsdorp, Western Cape Photo by Les Underhill
Habitat
The Brown-throated Martin is usually associated with water, occurring near streams, rivers, dams, open wetlands, estuaries, pans, and sewage works. It appears to prefer wetlands in relatively open habitats. It regularly forages far from water over dry land, occasionally even desert scrub. The Brown-throated Martin is found in all biomes in southern Africa. It is most common in the fynbos, Grassland, Succulent, and Nama biomes. The Brown-throated Martin is less common and more patchy in woodland environments.
Typical habitat provided by a small farm dam. Near Worcester, Western Cape Photo by Les Underhill
Behaviour
The Brown-throated Martin is a resident, nomad, and short-distance migrant. It is present throughout its range at all times of the year, but its local occurrence depends on the availability of suitable wetland habitats and nesting sites.
Brown-throated Martin Riparia paludicola Near Himeville, KwaZulu-Natal Photo by Pamela Kleiman
The Brown-throated Martin is usually gregarious. Groups are most often seen foraging over water or nearby open areas. Their flight is rather fluttering and less dashing than the flight of larger swallows. May also be seen perched on shoreline vegetation like reeds, tall grass, bushes or overhanging trees, as well as fences. They sometimes also sit on the ground, especially during windy or inclement weather. When not breeding, roosts communally at night on reeds, huddled together in small groups.
Brown-throated Martin Riparia paludicola Near Bethlehem, Free State Photo by Janet du Plooy
The Brown-throated Martin is primarily diurnal, but forages until after sunset, especially at aquatic insect or termite alate emergences. Frequently forages with other swallows and swifts over water, adjacent marshes and grassland, skimming insects from water surface or snapping them up in flight. During calm weather, the Brown-throated Martin typically forages within 3 m of the water surface. They also regularly forage away from water over nearby dryland habitats.
The diet of the Brown-throated Martin consists mostly of flying insects, including mosquitoes, flies, midges, ant and termite alates, small beetles, grasshoppers, and small dragonflies.
Brown-throated Martin Riparia paludicola Riversdale district, Western Cape Photo by Terry Terblanche
The breeding season varies regionally, probably depending on a combination of rainfall, abundance of food, and availability of nesting sites. In the Western Cape, most breeding takes place between September and December, although there are records from most months of the year. Further north and east, the Brown-throated Martin is recorded to breed over a longer period, with more records in the drier winter and spring.
Brown-throated Martin Riparia paludicola Msunduze River, KwaZulu-Natal Photo by Malcolm Robinson
The Brown-throated Martin nests in self-excavated burrows in sandy soil in vertical sandbanks along rivers, road cuttings or quarries. They may sometimes also use old, abandoned, or incomplete burrows of other bird species, including bee-eaters and Pied Starlings. They are mostly colonial nesters, but pairs are monogamous. The nest burrow is up to 90cm long and ends in a nest chamber. The nest itself is a pad of grass and other fine plant material with feathers. 2 to 4 plain white eggs are laid per clutch. Incubation is done mostly by the female and lasts up to 12 days. The nestling period takes a further 25 days or so, during which time the young are fed by both parents.
Brown-throated Martin Riparia paludicola Richtersveld National Park, Northern Cape Photo by John Todd
Further Resources
This species text is adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Janet du Plooy, John Todd, Les Underhill, Malcolm Robinson, Pamela Kleiman, Phillip Nieuwoudt , Ryan Tippett, and Terry Terblanche is acknowledged.
Recommended citation format: Tippett RM. 2026. Brown-throated Martin Riparia paludicola. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/05/26/brown-throated-martin-riparia-paludicola/
List of bird species in this format is available here.
Cover image of Cape Shoveler by Cobus Elstadt – Near Jeffreys Bay, Eastern Cape
The Cape Shoveler, being a type of duck, belongs to the Family ANATIDAE (Ducks, Geese, Swans, and Waterfowl). The family has a cosmopolitan distribution, occurring on all the world’s continents except Antarctica. These birds are adapted for swimming, floating on the water surface, and sometimes diving. They range from medium to very large and have robust, broad, and elongated bodies. They generally have broad and flattened bills with a curved nail at the tip of the upper mandible and most have lamellae inside their bills for filter feeding. Their necks are moderately to very long, and the wings are short and pointed, supported by strong wing muscles. The short, strong legs are set far back on the body, and the feet are webbed. The family contains around 174 species in 53 genera. 20 species have been recorded from southern Africa.
Identification
The Cape Shoveler is a medium-sized duck with a distinctive, long spatulate bill. The sexes differ slightly in plumage variation.
Cape ShovelerAnas smithii – Male Strandfontein Sewage Works, Western Cape Photo by Rene Navarro
The adult male has a pale greyish head with small, faint, dark streaks. The rest of the body varies from greyish-brown to dark brown with blackish mottling. The rump and upper tail coverts have a blue-green sheen. The tail is blackish-brown, with cinnamon outer tail feathers. In flight, it shows a pale blue forewing that is separated from the green speculum by a white wing bar (diagnostic). The bill is black, and the eyes are yellow. The legs and feet are also yellow but turn to orange when breeding.
Cape ShovelerAnas smithii – Female Stilbaai, Western Cape Photo by Johan van Rooyen
Females resemble the males but are paler and greyer overall. They also have darker heads, and the rump and tail coverts lack the glossy sheen. Females have duller blue forewings and a smaller, duller green speculum. Their bills are dark grey (not black) and they have dark brown (not yellow)eyes. The legs and feet are brownish. Juveniles are similar to the adult female but are duller.
The Cape Shoveler is not easily mistaken for other southern African duck species. It is however similar to the female Northern Shoveler (Anas clypeata) but the latter occors only as a rare vagrant in southern Africa. The female Northern Shoveler is paler and more rufous in colour. It has a larger bill with yellowish-brown to orange sides.
Cape ShovelerAnas smithii – Pair Strandfontein Sewage Works, Western Cape Photo by Daryl de Beer
Status and Distribution
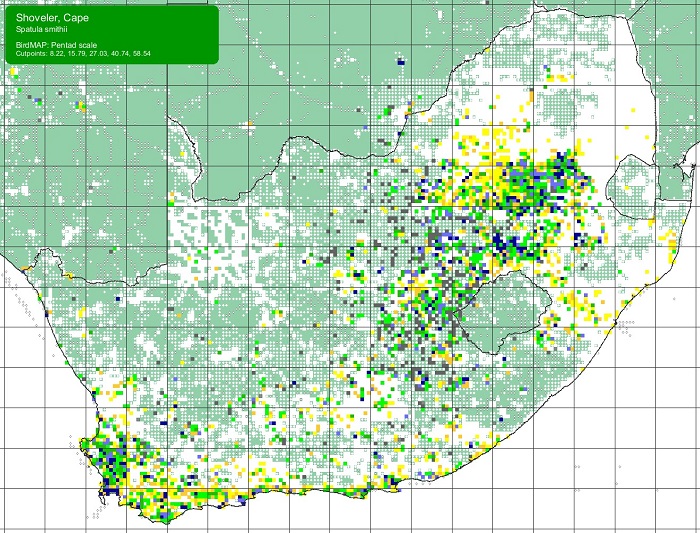
The Cape Shoveler is near-endemic to southern Africa where its status varies from rare to locally abundant. Its population is concentrated in South Africa. In South Africa, it is most abundant in the lowlands of the Western Cape, and in the highveld of the Free State, Mpumalanga, Gauteng and North West Provinces. Its population is otherwise scattered from the west coast through the Nama Karoo, Eastern Cape, and the coastal belt of KwaZulu-Natal. The Cape Shoveler is patchy or absent elsewhere.
It is an uncommon, occasional visitor to Lesotho, eSwatini (Swaziland). The Cape Shoveler is probably an irregular visitor to southern Mozambique and it is sparsely scattered throughout Namibia. It is locally common in south-eastern Botswana, but its range is scattered in northern Botswana, where it is absent from most of the Okavango Delta, but is common on the Makgadikgadi pan system in high rainfall years. The Cape Shoveler is irregular and scarce in western Zimbabwe and across the central plateau, mostly during the rainy season. Its range extends marginally beyond the region into southern Angola.
SABAP2 distribution map for Cape ShovelerAnas smithii – December 2024. Details for map interpretation can be found here.
The Cape Shoveler is not threatened. It appears to have extended its range and increased in numbers in the last 30 years. This is largely due to its fondness for artificial water bodies like dams and sewage ponds. It Additionally, it is not popular as a gamebird, nor is it favoured by aviculturalists. Hybridisation with introduced Mallards (Anas platyrhynchos) is a potential threat.
Cape ShovelerAnas smithii – Male Carnarvon district, Northern Cape Photo by Sybrand Venter
.
Habitat
The Cape Shoveler frequents a range of aquatic habitats but prefers shallow saline pans, seasonal marshes, tidal estuaries, sewage ponds, and shallow dams. It is also found at shallow bays and the upper reaches of large dams, as well as floodplains and subtropical pans. The Cape Shoveler favours highly alkaline and brackish waters. It avoids rivers, streams and acidic water bodies, such as occur in fynbos habitats of the Western Cape Province.
Ideal habitat at a shallow farm dam in the Nama Karoo. Carnarvon district, Northern Cape Photo by Ryan Tippett
Behaviour
The Cape Shoveler is most often encountered in pairs or small groups but may occasionally be found in flocks of up to 600 birds. In general, the Cape Shoveler keeps to itself but does sometimes form mixed flocks with other species. It is most likely to occur alongside the Cape Teal, which has similar habitat preferences.
Cape ShovelerAnas smithii – Male Strandfontein Sewage Works, Western Cape Photo by Felicity Ellmore
The movements of the Cape Shoveler are not well understood. Across their range birds may be migratory, nomadic or resident. Movements are known to occur between the winter and summer-rainfall regions. Nomadic movements are likely in response to food availability.
Cape ShovelerAnas smithii – Pair Rietvlei, Western Cape Photo by Sharon Stanton
When taking flight, the Cape Shoveler rises steeply with noisy wing-beats, reaching speeds up to 90 km/hr. In flight, the wings make a distinctive ‘whooping’ sound. The Cape Shoveler is secretive when in flightless moult. When disturbed they swim off low with only the head showing above the surface. The Cape Shoveler walks awkwardly on land. Groups spend much time loafing along the shoreline where they preen, rest or sleep, usually while standing in shallow water.
Cape ShovelerAnas smithii Nuwejaars Wetland Special Management Area, Western Cape Photo by Les Underhill
The Cape Shoveler feeds day or night and usually forages in shallow, open, nutrient-rich waters. They are primarily filter feeders, and most often forage by surface dabbling or with the head and neck submerged. They forage less often by upending or dabbling while walking. Cape Shovelers are also recorded to find food by ‘circle-swimming’ whereby a group of birds swim around in a circle, head to tail, filtering water stirred up by the bird ahead. The bulk of the Cape Shoveler’s diet comprises animal matter, including snails, insects, crustaceans, planktonic invertebrates, and tadpoles. Only around 30% of its diet consists of plant matter.
Cape ShovelerAnas smithii Pietermaritzburg, KwaZulu-Natal Photo by Malcolm Robinson
In the summer rainfall parts of its range, the Cape Shoveler breeds throughout the year with a small spring peak from July to September. Breeding in the winter rainfall Western Cape is more seasonal with a marked December peak and only a few breeding records in other months. The Cape Shoveler is a monogamous, solitary nester. Pair bonds are mostly seasonal, but may last for more than 1 season.
The nest is a grass-lined hollow in the ground ringed with down feathers. Nests are typically placed in dense vegetation, usually within 20m of the water. 5 to 12 cream coloured eggs are laid per clutch. Incubation is by the female only and takes 27 to 28 days. The young are precocial and are fully fledged by 8 weeks of age. The young are attended solely by the female.
Cape ShovelerAnas smithii – Female with young. Jongensfontein, Western Cape Photo by Gerald Gaigher
Further Resources
Species text adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Cobus Elstadt, Daryl de Beer, Felicity Ellmore, Gerald Gaigher, Gerald Wingate, Johan van Rooyen, Les Underhill, Malcolm Robinson, Rene Navarro, Ryan Tippett, Sharon Stanton, and Sybrand Venter is acknowledged.
Other common names: Kaapse slopeend (Afrikaans); iDada (Zulu); Canard de Smith (French); Kaplöffelente (German); Pato-trombeteiro do Cabo (Portuguese); Kaapse Slobeend (Dutch).
A list of bird species in this format is available here.
Recommended citation format: Tippett RM. 2026. Cape Shoveler Anas smithii. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/05/21/cape-shoveler-anas-smithii/
The African Oystercatchers on the coastline of Robben Island have been counted about 190 times since 2001. The earliest counts were of around 150 birds. The end of the blog will reveal what number was recorded on 17 May 2026.
Counting African Oystercatchers on Robben Island is not as easy as most people imagine it must be. The pair on the rocks in the foreground on the right are easy to spot, especially with the surf behind them. The second pair on the rocks among the cormorants and Kelp Gulls just right of centre are trickier.
Photo: Les Underhill
The oystercatcher of the left has its bright red bill tucked under a wing, and the red eye ring is just visible with binoculars, and its mate is partly obscured behind a rock.
Photo: Les Underhill
There are is a good variety of species on the shore. This Little Egret feeds in the rock pools at low tide, but during the spring high tide stands on a rock and waits for the opportunity to return to hunting.
Photo: Achot Ginosian
Spotted Thickknees roost in the vegetation above the high water line.
Photo: Achot Ginosian
Eurasian Whimbrels ought to be arriving on their breeding grounds in Siberia about now. We saw about 20. These are mostly young birds. Whimbrels do not start breeding until they are several years old, and it is pointless migrating 20,000 km there and back for nothing, so they stay put for the southern winter.
Photo: Achot Ginosian
African Penguins restarted breeding in Robben Island in 1984. In spite of the Apollo Sea oil spill of 1994 and the Treasure oil spill of 2000 knocking numbers downwards, they reached a peak abundance on the island in 2007. The number of breeding pairs now is about 10% of that peak.
Photo: Achot Ginosian
In the centre of the island, near the village is a large colony of Greater Crested Terns and also Hartlaub’s Gulls.
Photo: Achot Ginosian
After the start of the wet winter season, the Blue Stone Quarry, on the north side of the island, is full. The island has turned green after a long, dry, brown summer. There’s a handful of oystercatchers on the outcrop of rocks in the middle.
Photo: Achot Ginosian
The image below is a close up of the wing of the African Oystercatcher in flight above.
There is a distinct change in the shade of black between the inner primaries and the outer primary feathers. The newly grown inner primaries are dark black. The colour of the outer four primaries has faded from black to a brownish back. They have spent a year in the sun. Their tips are fraying; they have reached their “best before” date. The third primary from the end looks especially worn; the end of the feather has so few barbs that it is becoming transparent. In the gap between the new and old primaries is one that is halfway grown.
The process of feather replacement is energetically demanding. Adult African Oystercatchers start primary moult soon after the end of their midsummer breeding season, mostly in March and April. Moult continues at a slow pace through winter, taking five months, and finishes in spring, August and September. Within a couple of months, they will have their next breeding season, starting from November.
Photo: Les Underhill
The total for 17 May 2026 was 479 birds. This number is very much in line with recent counts of African Oystercatchers on Robben Island, and a three-fold increase since we started these counts 25 years ago.
The very first count made on the island was 40 African Oystercatchers on 18 August 1977, reported in Phil Hockey’s PhD thesis. The reasons for the massive increase include a huge reduction in human disturbance along the shoreline, and the invasion of the coastline with the Mediterranean Mussel, an alien species that grows faster than the indigenous mussels, and which grows higher up the intertidal zone, providing longer feeding opportunities for the oystercatchers on each tidal cycle. John Yeld wrote an article in GroupUp which provides useful background to the progress of the oystercatchers on Robben Island.
The Robben Island Museum has supported these oystercatcher surveys over the past 25 years, providing logistics on the island, and transport on the ferries to and from the island.
Achot Ginosian made a bird list. We finished on 33 species, which included of course … …
Photo: Chanan Weiss
… … the Chukar Partridge!
Here is the bird list! The species in red have links to the structured species text on the BDI website.
Rene found lots of these. Even though the Black-legged Golden Orb-web Spider is a large spider, up yo 30 mm, it is not as big as its English common name! Until 2002, this species was only recorded east of the Hottentots Holland Mountains. It then spread across the Cape Flats to the Cape Peninsula. It reached Robben Island a few years ago. Autumn is the time of the year when it is most abundant; it spins large webs between trees and shrubs.
We grabbed the opportunity presented by a calm Saturday morning to do bird ringing at the Strandfontein Sewage Works (relevant permissions having been obtained).
We were there well before sunrise to put up the mist nets.
The mist net poles are visible, but the net is not. Dieter is extracting a bird
The contrast between the primaries and secondaries of this Red-winged Starling make doing the moult score easier than it is for most species. The secondaries primaries are black and the primaries are “red” (that is not really “red”, that is “reddish brown”). It’s a female, because of the grey head. Compared with the pointed wings of long-distance migrants, the wings are quite rounded. The nine primaries that can be seen in the photograph are similar in size. There is a 10th primary on the outside of the wing (not visible here) which is about half as long as the ninth. In most passerines the 10th primary is tiny.
Red-winged Starlings moult in summer. So at some time during the past few months this bird has been through some kind of trauma, possibly a fight with another starling, and the tip of the fifth primary has been broken off. This damage won’t be patched up until the next moult, and meanwhile the aerodynamics are a bit compromised.
The bird was chill, and posed for photos before it was released.
Even an African Swamphen, out in bright sunlight on the edge of a reedbed, can be inconspicuous
We worked quite hard trying to catch this bird, both with a spring trap and with mist nets.
But alas, in spite of serious efforts, the final score was: Ringers 0 – Swamphen 1
There were lots of birds flying over. The ringer looks at them and asks the question: “Are they moulting?”
Look at the fifth primary of this Great White Pelican. That’s counting from the inside (the sixth counting from the outside). It is a bit shorter than its neighbours. It must be in moult. And the 10th primary, the outermost, looks like it is also in growth. Large birds, like pelicans, have complex moult patterns, and mostly do not moult all their primaries every year.
Looking at the right wing, the final three primaries of this African Sacred Ibis are in moult.
You will never see a Spur-winged Goose with growing primaries. Ducks, swans and geese do a catastrophic moult. The drop all their flight feathers simultaneously and become flightless (and vulnerable to predation). They regrow the feathers over a period of a few weeks. Coots, grebes, rails, and some other waterbirds also become flightless through their catastrophic moult. Most waterbirds can do at least some feeding during their moult. All penguin species do a catastrophic moult. But they cannot feed at all during moult because they lose their waterproofing, and therefore their insulation from the cold. They fatten up beforehand, come ashore for between two and four weeks and use their reserves to live, and to replace their feathers.
This is a close up of the bend in the wing of the Spur-winged Goose in the photo above. That’s the spur that gives the species its English common name.
Cover image of Grey-winged Francolin by Kate Webster – Mountain Zebra National Park, Eastern Cape
Francolins belong to the Family: PHASIANIDAE (Pheasants, Grouse, and allies). Other southern African representatives of this family include Spurfowls and Quails. They are generally plump, heavy-bodied gamebirds, with broad, relatively short wings and powerful legs. Many have a spur on each leg, while some, have reduced spurs or none at all. The bill is short and compact. Phasianids inhabit almost all terrestrial habitats, from Arctic and alpine tundra to grasslands, deserts, and savannas, to humid tropical rainforest. Most African species are cryptically patterned but many Asian and Australasian species are among the most spectacularly plumaged of all birds. The Phasianidae comprise 187 species from 54 Genera. 15 species are native to southern Africa.
Identification
The Grey-winged Francolin is a cryptically plumaged species, yet the feathers are beautifully and intricately patterned in black, white, brown and rufous. The sexes are alike in colouration, but males are larger than females.
Grey-winged FrancolinScleroptila afra Near Hartenbos, Western Cape Photo by Johan Heyns
In adults, the upper parts are grey-brown, with white feather shafts and with white or buff transverse barring. The head is warm brownish-buff, with dark speckling. The crown is streaked buff and blackish grey, and the ear coverts are brown. The nape and hind neck are dark grey, spotted white, and the pattern extends to the sides of the lower neck. The lower foreneck is pale cinnamon, and the feathers have distinct rufous patches. The underparts are paler and finely but irregularly barred black and off-white. Rufous primary coverts and primary bases are visible in flight. The bill is grey and heavy at the base, and noticeably decurved. The eyes are dark brown, and the legs and feet are dull greyish yellow. Males are equipped with sharp, robust leg spurs.
Grey-winged FrancolinScleroptila afra Swellendam district, Western Cape Photo by Johan van Rooyen
Immature and juvenile birds resemble adults, but carry duller plumage and white throat.
The Grey-winged Francolin is genetically closely allied to the Orange River Francolin (Scleroptila gutturalis), Red-winged Francolin (Scleroptila levaillantii), and Shelley’s Francolin (Scleroptila shelleyi), and it replaces these species in high altitude grasslands. The Grey-winged Francolin differs from other Scleroptila francolins in having more grey than red on the wings (red confined to the outer wing), and a grey-freckled, not white, or buff, throat. It differs from Shelley’s Francolin in having narrow barring on the underparts.
Status and Distribution
The Grey-winged Francolin is locally common. It is endemic to South Africa and Lesotho.
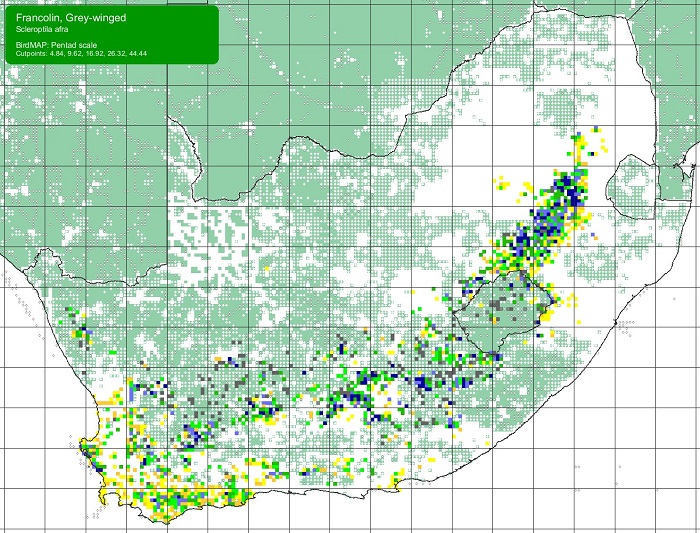
SABAP2 distribution map for Grey-winged FrancolinScleroptila afra – December 2024. Details for map interpretation can be found here.
The Grey-winged Francolin ranges from Mpumalanga, through the KwaZulu-Natal Drakensberg, Lesotho, the eastern Free State, and the highlands of the Eastern Cape westwards through the southern half of the Northern Cape to the southern Western Cape. The conspicuous gap in its distribution in the Great Karoo is probably due to sparse vegetation cover in that region. The Grey-winged Francolin is most common in the highland grasslands in the Drakensberg region. It is less numerous in the Northern and Western Cape.
The Grey-winged Francolin is partially sympatric with the Red-winged Francolin (Scleroptila levaillantii), but is generally found at higher altitudes. It occurs alongside the Cape Spurfowl (Pternistes capensis) in the succulent Karoo and fynbos areas of the W Cape.
Grey-winged FrancolinScleroptila afra Golden Gate Highlands National Park, Free State Photo by Terry Terblanche
The Grey-winged Francolin is not threatened. The historical distribution was probably similar to the current distribution. It is known to benefit from habitat modification due to moderate stock farming. Veld burning during early summer has the most detrimental influence on Grey-winged Francolins.
The Grey-winged Francolin is the only southern African francolin sufficiently abundant to sustain commercial wing-shooting. It has been hunted for recreation and commercially in the Eastern Cape for at least the past 120 years. This hunting is known to be both biologically sustainable and economically viable. Additionally, it provides landowners with an incentive to conserve grassland habitats. Hunted populations breed earlier than those that are not hunted, and are more outbred because of increased local movements following hunting.
Grey-winged FrancolinScleroptila afra Carnarvon district, Northern Cape Photo by Ryan Tippett
.
Habitat
The Grey-winged Francolin is typically found in mountainous areas in the Grassland, Nama Karoo, Succulent Karoo, and Fynbos biomes. It prefers ridge tops and shelves to steep slopes and valley bottoms. The Grey-winged Francolin is most numerous in montane grasslands, usually above 1800m. In the Succulent and Nama Karoo, it inhabits hilly and mountainous areas with sufficient grass and scrub cover. In the Fynbos biome, it occurs in grassy places in short fynbos, renosterbos, and strandveld down to sea level.
Montane grassland habitat. Golden Gate Highlands National Park, Free State Photo by Ryan TippettMountainous habitat in the Nama Karoo with sufficient grass and scrub cover. Carnarvon district, Northern Cape Photo by Ryan Tippett
Behaviour
The Grey-winged Francolin is resident with no evidence of seasonal movements. Breeding birds remain in the same general locations throughout their lives. However, young birds move between coveys often enough to ensure genetic interchange. Fluctuations in reporting rates reflect variations in conspicuousness linked to the breeding cycle.
Grey-winged FrancolinScleroptila afra Gamkaberg Nature Reserve, Western Cape Photo by John Todd
The presence of Grey-winged Francolins is usually revealed by territorial advertisement calls or when flushed. They are otherwise inconspicuous birds. The Grey-winged Francolin often warms up in the early morning on open ground, such as at road verges. Flushed birds fly fast and strongly, typically taking advantage of wind and steep ground to aid escape. Apart from this, they seldom take flight. The Grey-winged Francolin is most likely to run if disturbed (when vegetation is sparse), otherwise sits tight, flushing at the last moment. Birds roost on open ground, huddled together.
Orange River FrancolinScleroptila gutturalis West Coast National Park, Western Cape Photo by Felicity Ellmore
The diet of the Grey-winged Francolin varies seasonally. It mostly eats underground bulbs and corms of a range of small geophytes that it digs out with its bill. Invertebrates dominate the diet during the summer breeding season.
Grey-winged FrancolinScleroptila afra Carnarvon district, Northern Cape Photo by Ryan Tippett
The Grey-winged Francolin breeds during spring and summer throughout its range, with peak egg-laying between August and November. The breeding season is shorter in the winter rainfall Western Cape. The Grey-winged Francolin is monogamous. Pairs nest solitarily and they are probably territorial.
The nest is a scrape in the ground hidden under a grass tuft. It is lined with grass and occasionally feathers. 4 to 8 eggs are laid per clutch. The eggs are yellowish-brown with some brown and dark grey speckling. Incubation starts after the full clutch has been laid and last for up to 23 days. Incubation is performed soley by the female and she is guarded by the male.
The chicks are precocial, running and feeding almost immediately after hatching. Chicks stay with their parents throughout the breeding season. Breeding failures are mostly due to predation, primarily by small mammalian carnivores, crows and snakes. Other causes of nest failure include fires, human disturbance and trampling by sheep.
Grey-winged FrancolinScleroptila afra Near Bethal, Mpumalanga Photo by Vaino Prinsloo
Further Resources
Species text adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Felicity Ellmore, Johan Heyns, Johan van Rooyen, John Todd, Kate Webster, Keir Lynch, Ryan Tippett, Terry Terblanche, Tino Herselman, and Vaino Prinsloo is acknowledged.
Other common names: Greywing Francolin (Alt. English); Bergpatrys (Afrikaans); iNtendele (Zulu); Isakhwatsha (Xhosa); Khoale (South Sotho); Francolin à ailes grises (French); Grauflügelfrankolin (German); Francolim-d’asa-cinzenta (Portuguese); Grijsvleugel-frankolijn (Dutch).
A list of bird species in this format is available here.
Recommended citation format: Tippett RM. 2026. Grey-winged Francolin Scleroptila afra. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/05/11/grey-winged-francolin-scleroptila-afra/
Frogmore Estate is a suburb of Cape Town, a few kilometres north of Muizenberg, and bordering on Zandvlei. The Zandvlei Trust plays a key role in the conservation of this large wetland system. The fact that this a good bird ringing site is a tribute to their success.
On 9 May 2026, we had a line of five mist nets along a track through the reedbeds, and one mist net set over a bridge on the Keysers River. There was not the slightest breeze, so the nets hung like curtains. Ideal conditions.
Photo: Les Underhill
The clouds behind the mountain were the first sign of a big cold front due to arrive in about 15 hours, bringing heavy rain to the Western Cape. with severe weather warnings of floods, 100 km/h winds and dangerous waves at sea!
The list of birds we handled is below. There were 36 birds of 13 species, with none reaching double figures! (The species in red have a structured species account on the BDI website; there’s a full list of these species here.)
There were three retraps from previous ringing events at this site. We only have photographs of one of them, the Malachite Kingfisher below. One of the Levaillant’s Cisticolas had been ringed on 15 March last year, a little over a year ago. One of the Southern Double-collared Sunbird, a male, had been ringed as an adult on 23 March 2019. That’s a bit more than seven years since ringing, and because it was an adult already when ringed, it must be at least eight years old.
The two Malachite Kingfishers were in the hand at the same time. One was a retrap from 20 December last year, the day we held or end-of-year ringing event, for which there is a blog!
Photo: Achot GinosianPhoto: Achot Ginosian
The privilege of ringers is to see birds really close up.
Photo: Achot Ginosian
The bill of the Southern Marked Weaver looks a less formidable weapon that that of the Cape Weaver (see the photo below). This bird’s eye is turning red, so it is a male.
Photo: Achot Ginosian
Male Cape Weaver, with marie biscuit-coloured eye, and a longer bill than the Southern Masked Weaver.
Photo: Achot Ginosian
By May, primary moult has been complete for several months. Count the nine primaries of this Cape Weaver; it’s easiest to count from the outside in. The innermost was replaced first, and the outermost last. The process takes about four months to complete. The nine primaries of this bird still look new; they are not starting to fray at the tips. These feathers will have to do a lot of work before they get replaced just after the end of the upcoming breeding season, in spring. By then they will look worn out at the tips.
Lesser Swamp Warbler, in the mist net, and a few seconds later, in the hand in the ringer’s grip, with the head and neck held gently between the index finger and the middle finger, and with the bird’s body fitting snugly into the palm of the hand.
The mist net set over the Keysers River caught three Red-knobbed Coots.
Photo: Achot Ginosian
The red knobs, up close!
Photo: Achot Ginosian
Unlike most waterbirds, coots don’t have webbed feet.
The Keysers River is the largest waterway feeding into Zandvle. It drains a large area, from the watershed along the top ridge of the Constantiaberg, and the mountainside to the east of this. Then it collects all the stormwater from suburbs such as Tokai, Kirstenhof, Constantia and parts of Bergvliet and Retreat. It is likely to prove an important corridor for bird movement, especially of waterbirds.
Photo: Les Underhill
A list of dates for future ringing events is here.
There is a list of ringing expeditions that have blog reports similar to this here.
Cover image of Orange River Francolin by Lia Steen – Near Devon, Gauteng
Francolins belong to the Family: PHASIANIDAE (Pheasants, Grouse, and allies). Other southern African representatives of this family include Spurfowls and Quails. They are generally plump, heavy-bodied gamebirds, with broad, relatively short wings and powerful legs. Many have a spur on each leg, while some, have reduced spurs or none at all. The bill is short and compact. Phasianids inhabit almost all terrestrial habitats, from Arctic and alpine tundra to grasslands, deserts, and savannas, to humid tropical rainforest. Most African species are cryptically patterned but many Asian and Australasian species are among the most spectacularly plumaged of all birds. The Phasianidae comprise 187 species from 54 Genera. 15 species are native to southern Africa.
Identification
The Orange River Francolin is cryptically coloured with intricately patterned plumage. The sexes are alike in plumage coloration, but males are larger than females.
Orange River FrancolinScleroptila gutturalis Sasolburg, Free State Photo by Dawie Kleynhans
Adults are overall greyish-brown on the upper parts with white feather shafts, and pale buff central stripes and transverse bars. A narrow black-and-white band runs down the sides of the neck from behind eye. It ends on the sides of the lower neck and does not form a necklace. A black moustachial stripe extends as a second black-and-white band that does join as a necklace around white throat. The upper breast and flank feathers are buff-coloured, with large red-brown blotches. The belly feathers are also buff-coloured, with indistinct black-and-white barring that varies between races. In flight, the outer secondaries, primaries, and primary coverts are extensively rufous. The bill is strong, decurved, and grey, with a yellow base to the lower mandible. The eyes are brown, and the legs and feet are yellow. Male are equipped with sharp tarsal spurs.
Orange River FrancolinScleroptila gutturalis Near Balfour, Mpumalanga Photo by William Price
Immature and juvenile birds resemble the adults, but the black-and-white bands on the head and neck are less well defined, and their underparts are irregularly barred black and buff.
The Orange River Francolin is genetically closely allied to the Grey-winged Francolin (Scleroptila afra), Red-winged Francolin (Scleroptila levaillantii), and Shelley’s Francolin (Scleroptila shelleyi), and it replaces these species in arid grasslands. It is most likely to be confused with Shelley’s Francolin and Red-winged Francolin. The face pattern is most similar to that of Shelley’s Francolin, but in the latter, both dark facial stripes meet on the foreneck. The Orange River Francolin also lacks the black belly markings of Shelley’s Francolin and has a faster and higher-pitched call. The Red-winged Francolin lacks the black-and-white stripe behind the eye of the Orange River Francolin.
Orange River FrancolinScleroptila gutturalis Olienhout Game Reserve, Northern Cape Photo by Ryan Tippett
Status and Distribution
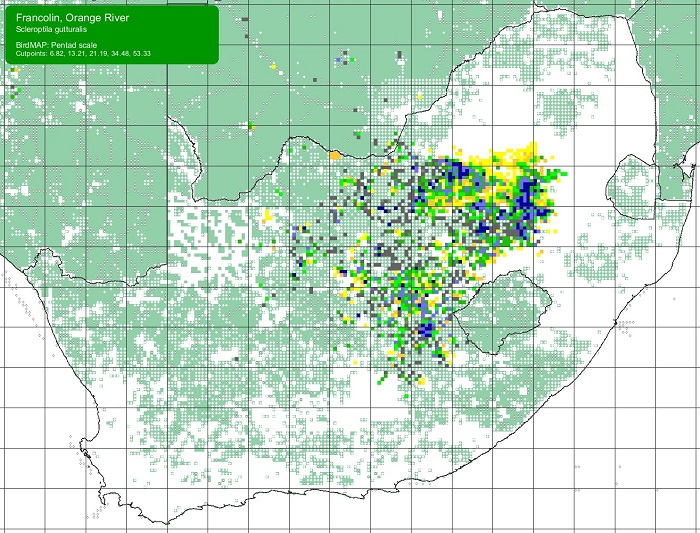
The Orange River Francolin is locally common in southern Africa. It has a curious and discontinuous distribution in north-east and southern Africa with several disjunct populations between Ethiopia, northern Kenya, and Uganda. It also has an anomalous southern African range, extending from the dry grasslands of central South Africa diagonally north-west through the sparse woodlands of the central Kalahari in Botswana, to northern Namibia and beyond into south-western Angola. In South Africa, the Orange River Francolin occurs in the north-eastern parts of the Northern Cape, much of the North West Province and Free State, and the extreme north-eastern parts of the Eastern Cape and across to the highveld of Gauteng and western Mpumalanga.
SABAP2 distribution map for Orange River FrancolinScleroptila gutturalis – December 2024. Details for map interpretation can be found here.
Overall, the Orange River Francolin is not threatened. However, it was apparently previously abundant and widespread throughout the Northern Cape. Since the 1930s, the Orange River Francolin has all but disappeared west of 22°E. The evidence and reasoning for this, though, are unclear. 2957 wild-shot birds were sold at the Kimberley market in 1906, but it is thought unlikely that this hunting pressure is the cause of consistently low numbers.
Reports from the early 1900s indicate that populations in South Africa were limited by annual grass burning just before the breeding season and again during the breeding season, by burning of any previously unburned patches. The Orange River Francolin is susceptible to habitat manipulation, particularly where poorly managed grazing results in degraded grasslands. Possible reasons for population decrease include the loss of habitat to agriculture and overgrazing. A few consecutive years of overgrazing, even on well-managed nature reserves, can lead to local extinctions.
Orange River FrancolinScleroptila gutturalis Suikerbosrand Nature Reserve, Gauteng Photo by William Price
.
Habitat
Dry grassland habitat. Near Petrusville, Northern Cape Photo by Ryan Tippett
The Orange River Francolin inhabits relatively arid grassland in either open or lightly wooded regions, typically on sandy or gravelly soils. In South Africa, it inhabits open grassland in the eastern and southern parts of its range, and wooded and bushy grasslands in the north and west. The Orange River Francolin prefers relatively flat or rolling terrain in South Africa, but favours hilly and mountainous terrain in Namibia. In Botswana, it typically prefers fossil river valleys. The Orange River Francolin is partial to short, sparse grasslands dominated by Themeda, Digitaria, Aristida, and Eragrostis species. It also forages at the edges of cultivated lands and fallow fields adjacent to its natural habitat.
Sparse, arid grassland in the southern Kalahari. Olienhout Game Reserve, Northern Cape Photo by Ryan Tippett
Behaviour
The Orange River Francolin is resident with no evidence of seasonal migration. However, they undertake considerable local movements during droughts and in response to habitat deterioration.
Orange River FrancolinScleroptila gutturalis Near Balfour, Mpumalanga Photo by William Price
The Orange River Francolin is usually found in pairs or small coveys of 3 to 5 birds, and occasionally in groups of up to 8. When approached, they tend to sit tight before scuttling away into thicker cover. If flushed, they gain considerable height before peeling off over trees, often increasing speed by flying downwind. They spend most of the day foraging or resting, and are constantly alert. Coveys regularly dustbathe around midday. Roost sites are invariably located in flat, open patches and are seldom used for more than two nights in a row.
Orange River FrancolinScleroptila gutturalis Near Balfour, Mpumalanga Photo by William Price
The Orange River Francolin forages by digging up underground food using its bill. Their diet comprises corms, bulbs (including Oxalis and Moraea spp), berries, seeds, flowers, fallen grain and some green shoots. They also consume a variety of insects, including bugs, beetles, ants, termites, grasshoppers and locust nymphs. Their diet is more plant-based in winter and supplemented with insects in summer.
Orange River FrancolinScleroptila gutturalis Suikerbosrand Nature Reserve, Gauteng Photo by William Price
The Orange River Francolin is recorded breeding during all months but mainly after early and late summer rains. It is a monogamous breeder and pairs nest solitarily. They are likely to be territorial. The nest is built entirely by the female. It is a simple scrape in the ground, usually hidden in dense grass, and lined with dry grass. Nearby vegetation is sometimes pulled over the nest to form a canopy.
Laying dates are variable, according to rainfall. 3 to 8 eggs are laid per clutch. The eggs range in colour from pale brownish to yellowish, sometimes with light brown speckles. Incubation starts after clutch completion and eggs hatch synchronously. The incubation period lasts up to 21 days and all incubating is done by the female. Newly hatched young are precocial and the young are cared for by both parents. They are brooded by the female at night until they are around 40 days old.
Orange River FrancolinScleroptila gutturalis Olienhout Game Reserve, Northern Cape Photo by Ryan Tippett
Further Resources
Species text adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Dawie Kleynhans, Lia Steen, Ryan Tippett, and William Price is acknowledged.
Other common names: Kalaharipatrys (Afrikaans); Francolin d’Archer (French); Rebhuhnfrankolin (German); Francolim-dourado (Portuguese); Archer-frankolijn (Dutch).
A list of bird species in this format is available here.
Recommended citation format: Tippett RM. 2026. Orange River Francolin Scleroptila gutturalis. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/04/30/orange-river-francolin-scleroptila-gutturalis/