On 14 to 19 June 2026 we made our second visit this year to the Nuwejaars Wetlands Special Management Area, the first visit have been made in April, with blog here. On this trip, we handled 149 birds of 34 species, as detailed in the table below.
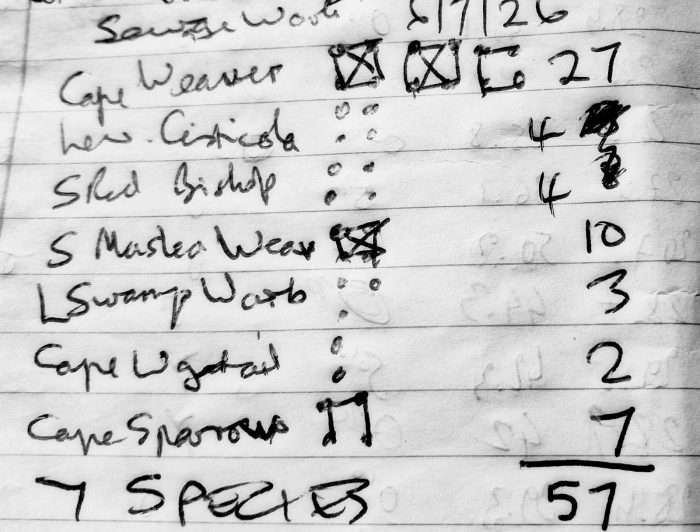
Multiple bird ringing expeditions to the Strandfontein Sewage Works have taken place since the one described in the blog for 24 May 2026. On that trip we ringed 27 Lesser Swamp Warblers, and we thought that was amazing. Subsequently, close on 200 Lesser Swamp Warblers have been ringed at the sewage works, with a maximum of 56 on 7 July.
Dieter Oschadleus is exploring lots of different ringing places at the sewage works for Lesser Swamp Warblers. The spot we chose this day was not particularly productive, and we handled only four!
We also caught a Red-knobbed Coot. Two fascinating things to look at with a coot in the hand are the red knobs, and the amazing feet:
Although we did not hit the jackpot from a bird ringing perspective (nine birds of four species), it was a great morning to be out-of-doors:
Sunrise, 20 July 2026. That’s a Yellow-billed Duck flying over Zeekoeivlei. The east face of Table Mountain is on the left and Devil’s Peak is on the right; the Saddle, in between is draped in cloud.
Spring is early, and the “nik nak lilies” are already putting on a good show. The Arum Lily Zantedeschia aethiopica is indigenous in southern Africa, but a dreadful invasive alien in many parts of the world, including Australia and New Zealand.
This excavation under the boundary fence is used by Cape Clawless Otters (see footprint) to move between Zeekoeivlei and the sewage works.
You will find a list of future bird ringing events here, and a list of past events with blogs here.
Cover image of Cardinal Woodpecker by Marna Buys – Magaliesberg Natural Area, Gauteng
Woodpeckers belong to the family PICIDAE. Members of this family are chiefly known for the characteristic ‘woodpecking’ behaviour that serves foraging, communication through drumming, and nest excavation. They have straight, strong bills that are reinforced at the base to cushion the brain, a stiff-pointed tail, and most possess zygodactyl toes. The tongue wraps around the rear of the skull and is anchored in one of the eye sockets or nostrils. The colours of many species are based on olive and brown, and some are pied, while others are boldly patterned in black, white, and red. Many have a crest or tufted feathers on their crowns. Woodpeckers have a mostly cosmopolitan distribution, although they are absent from Australasia, Madagascar, and Antarctica. They have diversified into every treed habitat on Earth, and most species live in forested or woodland habitats, although a few species have adapted to largely treeless environments. The family comprises 36 genera and 236 species. 10 species are known from southern Africa.
Identification
The Cardinal Woodpecker is the smallest woodpecker species in southern Africa, reaching a maximum length of 16cm and weighing up to 30 grams. Cardinal Woodpeckers are sexually dimorphic. The sexes differ in their head markings, and males are slightly larger than females, with longer wings and bills.
Cardinal WoodpeckerDendropicos fuscescens – Male Mokala National Park, Northern Cape Photo by Sybrand Venter
In adult males, the forehead to mid-crown is brown, and the hind crown and nape are red. A blackish-brown malar stripe extends from the base of the bill to the sides of the neck. The rest of the facial area and throat is white to greyish-white, sparsely and finely streaked dark brown. The upper parts are olive green with alternating bands of blackish brown and white barring. The tail is brown with yellowish barring and golden yellow feather shafts. The underparts are greyish-white with heavy brownish-black streaking. The bill is black and the tongue is long with a barbed tip. The eyes are dark reddish-brown and the legs and feet are greyish black.
Adult females resemble the males, but have a black hind crown and nape. Juveniles are duller and greyer than adults.
Cardinal WoodpeckerDendropicos fuscescens – Female Thanda Private Game Reserve, KwaZulu-Natal Photo by Ryan Tippett
The Cardinal Woodpecker is sometimes confused with the Golden-tailed Woodpecker (Campethera abingoni), which is the only other woodpecker in the region with streaked underparts. The Cardinal Woodpecker is much smaller and has a conspicuously barred (not spotted) back.
Cardinal WoodpeckerDendropicos fuscescens – Male Near Bela-Bela, Limpopo Photo by Ansie Dee Reis
Status and Distribution
The Cardinal Woodpecker is common throughout its range. It is confined to sub-Saharan Africa, from Gambia and Sudan down to South Africa. It is the most widespread woodpecker of the African savannas. The Cardinal Woodpecker is found throughout most of southern Africa and is only absent from treeless regions.
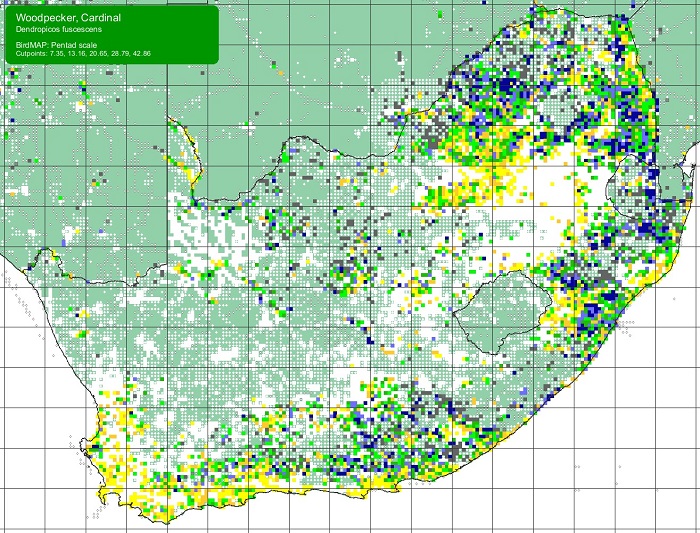
SABAP2 distribution map for Cardinal WoodpeckerDendropicos fuscescens – downloaded in December 2024. Details for map interpretation can be found here.
The Cardinal Woodpecker is not threatened. Its range has undoubtedly contracted in areas where woodland has been cleared for agriculture. However, its range has also expanded into areas where alien trees have been planted, where it previously did not occur.
Habitat
Typical arid savanna habitat. Mokala National Park, Northern Cape Photo by Karis Daniel
The Cardinal Woodpecker frequents a wide variety of woodland and savanna habitats, from the edges of deserts to the fringes of forests. It inhabits any woodland, includingthornveld, broad-leaved woodland and savanna. In desert and semi-desert areas, as well as fynbos and grassland regions, it is restricted to riparian tree galleries or introduced trees around settlements and farmsteads. It regularly enters villages and suburban environments. The Cardinal Woodpecker avoids the interior of evergreen forests, except where fragmented or regenerating. It is also absent from extensive monocultures of planted timber.
Subtropical woodland Habitat. Mkhuze Game Reserve, KwaZulu-Natal Photo by Colin Summersgill
Behaviour
The Cardinal Woodpecker is resident and sedentary. It is most often found in pairs, less often solitarily, and seldom in family groups except immediately after breeding. Pairs remain together and are territorial year-round. The Cardinal Woodpecker’s presence is usually given away by the tapping sound they make while foraging, or by its distinctive call.
Cardinal WoodpeckerDendropicos fuscescens Tembe Elephant Park, KwaZulu-Natal Photo by Phillip Nieuwoudt
Often forages on thin, mainly dead branches and stems of trees and shrubs that are not used by other woodpeckers. Cardinal Woodpeckers frequently join mixed-species foraging flocks, in which they may occur alongside other woodpeckers. In response to danger, will move out of view behind a branch. In territorial disputes, rivals call frequently, sway heads from side to side, raise crest feathers, flick wings open and closed, and pursue each other. At night, roosts solitarily in a hole in a tree, favouring holes with much larger entrances than nesting holes. Roost sites are reused nightly. Their flight is typically undulating, like most woodpeckers.
Cardinal WoodpeckerDendropicos fuscescens Shelley Beach, KwaZulu-Natal Photo by Lia Steen
This species is agile and dexterous while foraging, often hanging under branches and turning to peck open a site from different angles. It pecks open cavities and uses its barbed tongue to extract the larvae and pupae of beetles and other insects, including large caterpillars. Some prey, such as ants and spiders, are gleaned from bark and plant stems. It occasionally hawks termite alates and eats occasional small amounts of fruit. They also break into seed pods to feed on wasp larvae.
Cardinal WoodpeckerDendropicos fuscescens Ukhahlamba Drakensberg Park, KwaZulu-Natal Photo by Andre Harmse
The Cardinal Woodpecker is a monogamous, solitary nester. The nest is a hole excavated by both sexes, taking 13-21 days to complete. It is usually located on the underside of a dead, sloping stem up to 4 m above the ground. The nest cavity is up to 250 mm deep, with an oval-shaped entrance. Nest holes are seldom reused.
Cardinal WoodpeckerDendropicos fuscescens Albert Falls, KwaZulu-Natal Photo by Colin Summersgill
Eggs are mostly laid from July to December. 1 to 3 glossy white, oval-shaped eggs are laid per clutch on a bed of dry wood chips at 1-day intervals. Incubation starts at clutch completion and lasts up to 13 days. Both sexes share incubation duties. Eggshells and unhatched eggs not removed. Newly hatched young are altricial and are brooded and fed by both parents. The nestling period takes up to 27 days. Juvenile birds disperse within 2 months of fledging. Cardinal Woodpeckers are single-brooded.
Broods are frequently parasitised by honeyguides, including Lesser Honeyguide (Indicator minor), Greater Honeyguide (Indicator indicator), and Scaly-throated Honeyguide (Indicator variegatus).
Cardinal WoodpeckerDendropicos fuscescens Near Klerksdorp, North West Photo by Tony Archer
Further Resources
This species text is adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Christopher Willis, Colin Summersgill, Karis Daniel, Ryan Tippett, and Sybrand Venter is acknowledged.
Other common names: Kardinaalspeg (Afrikaans); iNqondaqonda (Zulu); Isinqolamthi (Xhosa); Pic cardinal (French); Kardinalspecht (German); Kardinaalspecht (Dutch); Pica-pau-cardeal (Portuguese).
Recommended citation format: Tippett RM. 2026. Cardinal Woodpecker Dendropicos fuscescens. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/07/17/cardinal-woodpecker-dendropicos-fuscescens/
A total of 141 species has been recorded for SABAP2 pentad 3340_2015, the one into which Ouberg Private Nature Reserve falls. That list is based on 29 full protocol checklists. There is one extra species recorded only on an ad hoc protocol checklist, making 142 species. We ringed 45 of these species, including some of the species that are seldom recorded, and have low reporting rates.
The list below is in reporting rate order. from smallest to largest. The reporting rate is the percentage of the 29 full protocol SABAP2 checklists for the pentad on which the species was recorded. A full protocol checklist is based on at least two hours of intensive searching for as many species as possible within the pentad, trying to visit as many of the habitats as feasible. We didn’t take a photo of some of the species, especially the ones with the largest reporting rates!
Dates of future ringing events from August to November 2026 are at the very end of this blog.
(Species in red like this have structured species texts on the BDI website. Click on the name, and you get taken to a page with lots of photos, an identification guide, the SABAP2 distribution map, and habitat photos. A full list of species with structured texts is here.)
The white tail spots of the Large Rock Martin look like this. They are conspicuous in flight when the bird is flying away from you and banks to make a sharp turn.
This White-backed Mousebird was the first bird handled on this expedition. Even though the pink-red legs and the white back are not visible, it is a White-backed Mousebird because the base of the bill is blueish white, and the tip is a dark grey-black.
The Acacia Pied Barbet’s reporting rate, 93%, means that it was recorded on 27 out of 29 SABAP2 checklists. This high reporting rate is largely explained by its loud and characteristic “toy-trumpet” call; it is not that abundant. It is dangerous to compare reporting rates between species solely from the perspective of relative abundance. Conspicuousness needs to be factored in as well. Reporting rates can be compared between pentads for a single species, but care needs to be taken doing that as well!
The species listed below all had high reporting rates, but there is only a photo of the Cape Robin-Chat!
Altogether, we handled 169 birds of 45 species at Ouberg! It is a very special place.
Artwork: Joël Daniel-Roux
There is a great variety of ringing sites at Ouberg: around the farmhouse, at various points in gaps in the trees and shrubs along the river courses, in the “bee area”, and even out in the open karoo scrubland. This diversity of ringing sites (and associated habitats) is reflected in the number of different species in the table below: 45 species. Only four species reached double figures: Cape Weaver (33), White-backed Mousebird (20), Cape Sparrow (12) and Cape White-eye (10). There were 46 retraps!
Species
Count
Acacia Pied Barbet
1
Barn Owl
1
Bar-throated Apalis
5
Black-headed Canary
1
Bokmakierie
2
Cape Penduline-Tit
1
Cape Batis
3
Cape Bulbul
2
Cape Robin-Chat
6
Cape Sparrow
12
Ring-necked Dove
1
Cape Wagtail
1
Cape Weaver
33
Cape White-eye
10
Cardinal Woodpecker
1
Chestnut-vented Warbler
6
Southern Fiscal
2
Common Waxbill
1
Fairy Flycatcher
2
Familiar Chat
1
Fiery-necked Nightjar
1
Fiscal Flycatcher
1
Greater Honeyguide
1
Grey Tit
1
Grey-backed Cisticola
2
Karoo Prinia
8
Karoo Scrub-Robin
6
Large-billed Lark
1
Layard’s Warbler
1
Lesser Honeyguide
2
Long-billed Crombec
1
Malachite Sunbird
3
Namaqua Warbler
1
Olive Thrush
1
Pririt Batis
2
Rock Martin
1
Rufous-eared Warbler
1
Southern Boubou
2
Southern Double-collared Sunbird
2
Southern Masked-Weaver
9
Southern Tchagra
3
Speckled Mousebird
5
White-backed Mousebird
20
White-throated Canary
1
Yellow-bellied Eremomela
1
Totals: Species : 45
169 birds
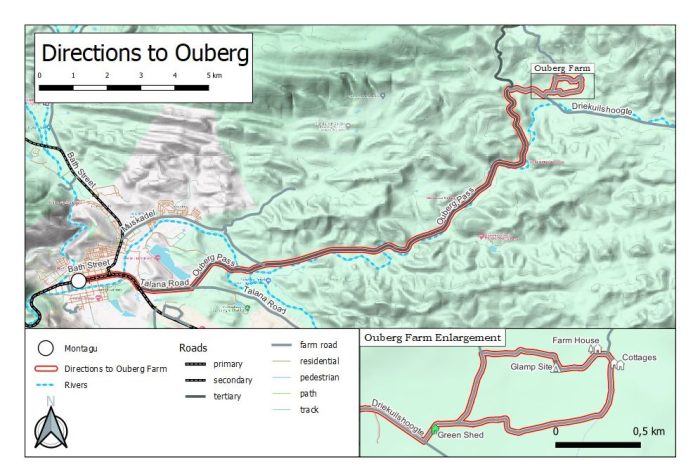
The next two ringing events at Ouberg Private Nature Reserve are from 5 to 10 September and from 18 to 23 October. If you would like to join one (or both!) of these, contact Sue Gie. The Ouberg Private Nature Reserve is about 20 km northeast of the town of Montagu in the Western Cape. See the map below!
There are also other ringing events on the horizon. 15 to 20 August at Nuwejaars Wetlands Special Management Area near Bredasdorp. There are two Open Days at Grotto Bay Private Nature Reserve, the first is on 12 September and the second is on 3 October. Email ring@thebdi.org for more information.
And most important of all is a Ringers’ Symposium at Nuwejaars Wetlands Special Management Area; the core dates for the symposium, with presentations are 8 to 10 November, with bird ringing events on the two days before and the two days afterwards, so the full symposium period is 6 to 12 November. The extended dates are set up with people who are traveling a long distance in mind.
Cover image of Red-capped Lark Lark by Ryan Tippett – Carnarvon district, Northern Cape
Larks are passerine birds of the family ALAUDIDAE. The larks are a group of mostly Old World passerine birds. Most species are found in Africa, followed by Asia and Europe with two species occurring in North America and one species in Australia. Habitats vary, but many are characteristic of open, dry regions. The family contains 21 genera and 100 species.
Identification
The Red-capped Lark is an attractive, slender, medium-sized lark with distinctive plumage, rendering it unlike any other southern African lark species. The sexes are alike, but males are slightly larger than females.
Red-capped LarkCalandrella cinerea Pienaarsrivier, Limpopo Photo by Ansie Dee Reis
Adult birds have very distinctive rufous patches on the sides of the chest as well as a rufous crown with a short erectile crest that is normally held flat. The crest is usually only raised when anxious or hot. The underparts are variable but are usually white, sometimes with buffy flanks. The lores and supercilium are white, and the ear coverts are whitish, with fine brown streaks. The nape, sides and back of the neck are brown. The mantle and back are pale to mid-brown; the mantle is finely streaked darker brown, while the back is more heavily marked. The rump is reddish brown with rufous upper tail coverts. The tail is dark brown with white outer tail feathers.
The Red-capped Lark is relatively long-winged, and the flight feathers are dark brown. Flight is pipit-like. The black bill is fairly small and slender. The eyes are brown, and the legs and feet are dark brown to black.
Red-capped LarkCalandrella cinerea Near Swellendam, Western Cape Photo by Johan van Rooyen
Juveniles resemble adults but carry darker upper parts with dark brown feather centres and buff margins with whitish tips. Juveniles also show a rufous-buff wash on the breast with some dark brown spots. The belly is creamy-buff (not white).
The Red-capped Lark is not easily mistaken for other species in southern Africa.
Red-capped LarkCalandrella cinerea Near Worcester, Western Cape Photo by Karis DanielRed-capped LarkCalandrella cinerea Bloemfontein, Free State Photo by Dawie de Swardt
Status and Distribution
The Red-capped Lark is common and locally abundant in South Africa but is less common in neighbouring countries.
It ranges from southern Kenya and eastern DRC south to southern Africa, with an isolated population in Nigeria. The Red-capped Lark is widespread in southern Africa, but is thinly distributed in much of Namibia and Botswana, and rare at low altitudes in Zimbabwe and Mozambique.
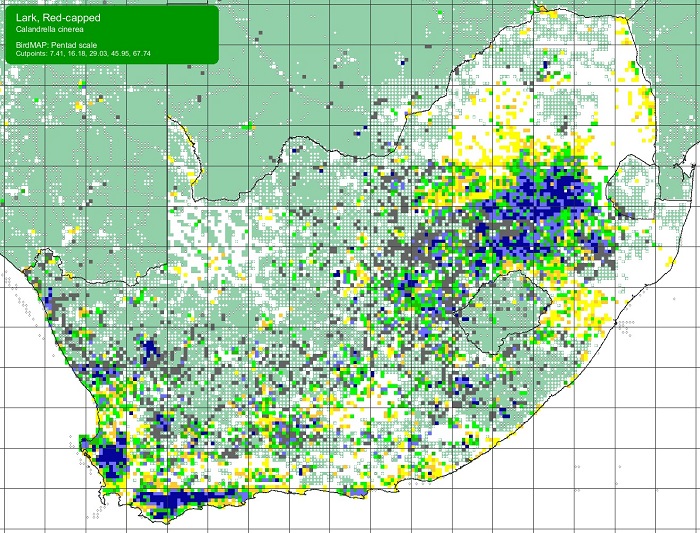
SABAP2 distribution map for Red-capped LarkCalandrella cinerea – July 2024. Details for map interpretation can be found here.
The Red-capped Lark is not threatened. It is well represented in protected areas and has adapted well to agricultural landscapes.
Red-capped LarkCalandrella cinerea Near Klerksdorp, North West Photo by Tony Archer
Habitat
Habitat near Greytown, KwaZulu-Natal. Photo by Colin Summersgill
The Red-capped Lark inhabit open grassland, bare ground in sparsely vegetated dwarf shrubland and the edges of wetlands. It also frequents short salt-marsh vegetation at coastal lagoons. It favours recently harvested or ploughed fields as well as short pastures. In the montane grasslands of eastern South Africa, it favours annually burned fields and heavily grazed grassland. The Red-capped Lark is frequently seen on roads and along road verges, especially after rain.
Habitat near Thaba-Tseka, Lesotho. Photo by Colin Summersgill
Behaviour
The Red-capped Lark may be resident, a partial migrant, or nomadic depending on the region and rainfall. Its movements are not well known. The Red-capped Lark is largely sedentary in some areas, but with more or less regular movements into both breeding and non-breeding areas, especially in mesic areas. There is a clear influx into Zimbabwe and eastern South Africa during winter, with birds moving north and west in summer. The Red-capped Lark is nomadic in the drier parts of its range.
Red-capped LarkCalandrella cinerea Near Senekal, Free State Photo by Johan van Rooyen
The Red-capped Lark is usually found in flocks of 5 to 20 birds. Under favourable conditions, flocks can number into the hundreds, or rarely thousands of birds.
Drinks water regularly. At least some birds fly to bare areas after sunset to roost. When disturbed, flies away low, showing white outer tail, then drops to ground and runs, or towers high into air and either circles to land nearby or flies a considerable distance.
Red-capped LarkCalandrella cinerea Carnarvon Dam, Northern Cape Photo by Ryan Tippett
Red-capped Larks forage by walking on the ground, occasionally running, pecking food from the soil surface or gleaning from foliage of grasses or shrubs. Breaks up antelope and sheep droppings with its bill to obtain seeds, and probes in cattle dung for insect larvae. Forages on burnt ground immediately after fire, but keeps away from the flame front.
Feeds on snouted harvester termite (Trinervitermes sp) workers when mounds have been broken open by aardvark.
Occasionally hawks moths or emerging termite alates aerially. Eats invertebrates and seeds. Stomach contents dominated by seeds, apart from scale insects, which feed on roots but emerge after rains; other insects important only when feeding chicks. Seeds incl grasses, sedges, shrubs, and forbs. Insect food incl ants, mantids, termites, ad and larval Lepidoptera, bugs, scale insects, grasshoppers, wasps, weevils and snails. Stomachs often contain grit, apparently to aid digestion.
Red-capped LarkCalandrella cinerea Near Frankfort, Free State Photo by Lance Robinson
The Red-capped Lark
Monogamous. Solitary nester, but breeding densities in the south of its range up to 2-4 pairs/ha; nests sometimes only 25 m apart. Territorial; in W Cape, territory size ca 1 ha. Occasionally chases other spp from nest site. In pipit-like aerial display, male rises almost vertically into air to ca 50 m, then flies into wind, flapping slowly, rising and dipping while singing; remains in air up to 10 min, gradually descending to 30-40 m, then closes wings and drops, either landing or flying up again to continue display. Males sometimes resort to fights on ground; threat display a crouched posture, with tail fanned and wings raised and quivering.
Red-capped LarkCalandrella cinerea Near Kimberly, Free State Photo by Dawie de SwardtRed-capped LarkCalandrella cinerea Carnarvon district, Northern Cape Photo by Ryan Tippett
Further Resources
Species text for Red-capped Lark in the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Ansie Dee Reis, Colin Summersgill, Dawie de Swardt, Johan van Rooyen, Karis Daniel, Lance Robinson, Ryan Tippett, Tino Herselman, and Tony Archer is acknowledged.
Virtual Museum (BirdPix > Search VM > By Scientific or Common Name).
Other common names: Rooikoplewerik (Afrikaans); umNtoli (Zulu); Intutyane (Xhosa); Rotscheitellerche (German); Alouette cendrille (French); Roodkapleeuwerik (Dutch); Cotovia-de-barrete-vermelho (Portuguese).
List of bird species in this format is available here.
Recommended citation format: Tippett RM 2026. Red-capped LarkCalandrella cinerea. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/07/15/red-capped-lark-calandrella-cinerea/
We were bird ringing near the Maskam Mountain, Vanrhynsdorp, from 3 to 7 July. We were based at Botuin Cottages. We ringed in the Botuin garden, the Lark Site close to the Maskam, and at the Vanrhynsdorp Sewage Works.
Friday 3 July
There was no wind from lunchtime. when we arrived at Botuin, until we went to bed.
In the afternoon, we had mist nets dotted around the Botuin garden, including in the olive grove …
Pride of place goes to the Lesser Honeyguide. This is a species not only ringed here for the first time, it is also the first atlas project record of the species in SABAP2 pentad 3135_1845 which has 264 full protocol checklists, and 182 species recorded to date.
The photo below shows one of the diagnostic features of the Lesser Honeyguide: the white feathers in the tail.
Just before lunchtime, we did a recce to the Lark Site!
It turned out not only to be a mist netting recce. The important take-home message which we discovered is this: “IF YOU WANT TO SEE THE WILDFLOWERS THIS YEAR, GO SOON!!” There is still some accommodation available at Botuin. Book today.
The Namaqua Daisies were stunning.
Sunday 5 July
Another day without wind.
We were at the Lark Site before first light, and had mist nets in place well before sunrise. A 07h13, the cliffs of the Maskam Mountain glow in the dawn light.
The Cape Clapper Larks were already displaying vigorously. But it will be another 30 minutes before the rays of the sun reach the cliffs.
The line of mist nets in the middle distance is still hard to spot. It is close to the fence line on the right hand side of the photo.
07h49. There is now real sun on the cliffs. but not yet on the slopes. This is a place with awe-inspiring presence.
The mist nets are more or less visible, directly below the right edge of the sunlit cliff.
From the perspective of numbers, we hit the jackpot. Just after sunrise, there was a big movement of Southern Red Bishops. and we handled 50! We handled 78 birds of 12 species. The seven species in bold face type in the list below were new, so the species total for the trip grew to 24:
In spite of this spot being called the Lark Site, we only caught a single lark: Cape Clapper Lark. It was our first bird, mistnetted before the sun was up.
Cape Clapper Lark
There is rather little SAFRING data for the Cape Clapper Lark. According to the SAFRING website, there are only five previous ringing records. So this is the sixth!
Achot took this photo of a Cape Clapper Lark in mid-display!
Rufous-eared Warbler
The urgency of coming to see the flowers is underlined by this male Southern Red Bishop. It is already coming into its red and black breeding plumage.
There is doucmentary about the Maskam, made for “people in cities who drive through this region” and see it as an inconvenient waste of time as they travel to their destination. The link to the 29-minute video is here. Salome is interviewed and Botuin Guesthouse gets a special mention. A good starting point is this article in the Daily Maverick: Exploring Maskam: A documentary about love, culture, and the mountain that unites.
Monday 6 July
Today it was the turn of the Vanrhynsdorp Sewage Works for a visit. The reflections of the clouds in the settling pond indicates that there is no wind again today.
It might be sunny but it is not as warm as Achot makes it look. The Maskam is a landmark that keeps a watchful eye on us.
In the afternoon we went back to the Lark Site, set up mist nets in the afternoon for a couple of hours, and then furled them overnight. This is a different spot, 500m or so east of where we were a couple of days ago:
It’s late afternoon, and the daisies have packed up for the day. They will open again tomorrow morning, once the temperature is warm enough.
Species of the afternoon was Cape Penduline Tit. It weighed in at 7.9g.
Maskam Mountain late on Monday afternoon.
The ringing total for the afternoon was eight birds: two Southern Red Bishops, two Cape Weavers, one Southern Masked Weaver, one Grey-backed Cisticola, one White-throated Canary and one Cape Penduline Tit. Species total 29.
Tuesday 7 July
… and we were back at Maskam Mountain early on Tuesday morning …
The morning was misty, and it was absolutely calm from the time we started until we left Botuin at noon. What a privilege to have five windless days in a row!
We caught our first and only Cape Bunting.
The long back claw gives this lark its common name: Spike-heeled Lark.
The white tail tips are conspicuous in flight. The scalloped patterning of many of the feathers, especially the coverts on the wing and tail, suggest that this is a young Spike-heeled Lark, most likely from last year’s breeding season.
Our final new species for the trip was this Karoo Lark. We handled 18 birds in a couple of hours, of 12 species. Three species were new, and that brought the species total to 32.
So the final total was 206 birds handled of 32 species.
… and to finish, here are a few of Achot’s photos of other good things which we were lucky to observe:
Blue Cranes are a newish arrival in the Vanrhynsdorp district. Numbers have been increasing over the past two decades. In the early years of SABAP2, which started in 2007, observations of Blue Cranes generated “Out of Range Forms”.
On this trip, we caught only two larks in total at the Lark Site! One of the species we missed out on was this Large-billed Lark.
This was one of a group of Bat-eared Foxes regularly seen along the road to the Lark Site.
These suricates have den in the Lark Site.
With the Maskam Mountains conspicuously dominating the background to so many of the photos above, now is the time to watch the video. It is in Afrikaans, with excellent English subitles.
Our next visit to Botuin is soon! It is from Tuesday 21 July to Sunday 26 July. There is still some accommodation available. Contact Les for further information.
Cover image of Cape Grassbird by Ryan Tippett – Ithala Game Reserve, KwaZulu-Natal
The Cape Grassbird belongs to the Family MACROSPHENIDAE (African Warblers). This is a recently recognised collection of species from the once vast family Sylviidae. The family is confined to sub-Saharan Africa. Macrosphenidae comprises 18 species from 6 genera.
Identification
Cape GrassbirdSphenoeacus afer Near Impendle, KwaZulu-Natal Photo by Colin Summersgill
The Cape Grassbird is not sexually dimorphic (the sexes are alike).
In adults, the forehead, crown and nape are bright, rusty red with black streaks on the hind crown and nape. The lores and the short supercilium are buffy-white, and the ear coverts are rufous. They have a pair of narrow black moustachial and malar stripes on the face and throat, separated by a broader white sub-moustachial stripe. The rest of the chin and throat are white. The rest of the underparts are off-white and the breast is mottled blackish brown. The flanks and undertail coverts carry broad, blackish streaks.
The back, mantle and sides of the neck are mottled black and buff. The rump and upper tail coverts are tawny, and the rump is sometimes streaked black. The tail is rusty red, long and strongly graduated with a pointed tip. The bill is grey-black, and the legs and feet vary from brown to grey. The eyes are brown with a buffy-white eye ring and an indistinct black outer ring.
Cape GrassbirdSphenoeacus afer Near Fort Nottingham, KwaZulu-Natal Photo by Colin Summersgill
Juveniles are like the adults but are duller, with black streaks on the crown. The head, mantle and underparts are also paler and the flanks are lightly streaked.
The Cape Grassbird resembles the similarly sized Moustached Grass Warbler (Melocichla mentalis). The Cape Grassbird has streaked (not plain) upper parts and a narrow, rufous (not broad, olive-brown) tail with a pointed (not rounded) tail tip.
Status and Distribution
The Cape Grassbird is a locally common endemic. It occurs along the escarpment and at higher altitudes in the North West and Limpopo Provinces, Gauteng, Mpumalanga, eSwatini (Swaziland), Lesotho, and KwaZulu-Natal, west through the Eastern Cape to the Western Cape. There is also an isolated population in the highlands of eastern Zimbabwe and adjacent Mozambique. The Cape Grassbird is absent from drier regions such as the central Karoo, Namibia, and Botswana.
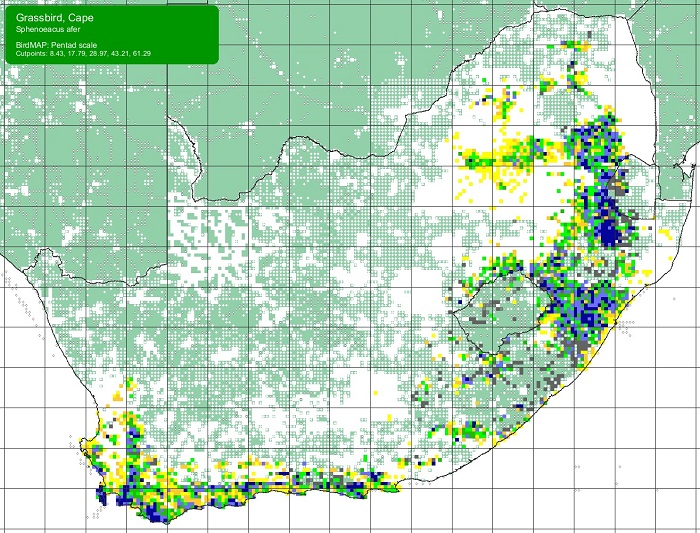
SABAP2 distribution map for Cape GrassbirdSphenoeacus afer – December 2024. Details for map interpretation can be found here.
The range of the Cape Grassbird is not known to have altered historically, although its abundance in certain regions is likely to have declined. It is not threatened, but has been negatively affected in the Western Cape by the invasion of fynbos by alien Acacia spp. The Cape Grassbird does not adapt to alien vegetation and abandons areas that become heavily infested. Its range in the Eastern Cape has also been impacted by poor veld management practices that have reduced the cover of rank grasses. In a number of areas, urban development, agriculture, and forestry have further reduced available habitat.
Cape GrassbirdSphenoeacus afer Near Swellendam, Western Cape Photo by Attie van Aarde
Habitat
The Cape Grassbird favours rank vegetation with long grasses, restios or ferns, and tangled scrub. It also lives in hilly grasslands with scattered bushes and boulders, and in rank vegetation along streams and at the edges of montane forest. In the Fynbos biome, it inhabits low-to medium-height fynbos dominated by restios. The Cape Grassbird avoids areas in which the woody component of the vegetation becomes too high or too dense. It occurs up to altitudes of 2300 m above sea level.
Habitat at Golden Gate Highlands National Park, Free State Photo by Ryan Tippett
Behaviour
The Cape Grassbird is resident and sedentary and is not known to undertake seasonal or altitudinal migrations. Seasonal differences in reporting rates are attributed to increased conspicuousness when singing during the breeding season.
Cape GrassbirdSphenoeacus afer Near Boston, KwaZulu-Natal Photo by Malcolm Robinson
The Cape Grassbird tends to skulk in dense vegetation, but its habit of sunbathing or singing loudly and distinctively from an exposed perch in the morning and late afternoon makes it unlikely to be overlooked or misidentified. It is unobtrusive when not singing. The flight appears laboured and clumsy, but is nevertheless fast and direct. The Cape Grassbird seldom flies far before dropping into vegetation.
Cape GrassbirdSphenoeacus afer Near Estcourt, KwaZulu-Natal Photo by Colin Summersgill
The Cape Grassbird searches for food on the ground, usually beneath vegetation. It is primarily an insectivore that consumes a variety of small insects, including beetles, grasshoppers, and caterpillars. It is also recorded feeding on small seeds and fruits.
Cape GrassbirdSphenoeacus afer Near Suurbraak, Western Cape Photo by Alan Manson
The Cape Grassbird breeds in early spring through to summer (July to December) in the Western Cape, and in summer (September to March) across the rest of its range. It is is a monogamous, solitary nester, and pairs are territorial.
The nest is a well-concealed cup of leaves, twigs and grass blades, lined with finer grass stems and other plant material. It is built entirely by the female. Nests are placed up to 400 mm above the ground in a tuft of grass or hidden in tangled growth. 2 to 3 eggs are laid per clutch at 1 day intervals. The eggs are white or pale grey, with slate-coloured dots. The incubation period takes up to 18 days and is done entirely by the female. The nestling period lasts further 14 to 16 days and the young are fed and cared for by both parents. Cape Grassbirds are sometimes double-brooded.
Cape GrassbirdSphenoeacus afer L’Agulhas, Western Cape Photo by Alan Manson
Further Resources
This species text is adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Alan Manson, Attie van Aarde, Colin Summersgill, Malcolm Robinson, and Ryan Tippett is acknowledged.
Other common names: Grassbird, Cape Grass-Warbler (Alt. English); Grasvoël (Afrikaans); Itshitshi, Udwetya (Xhosa); Sphénoèque du Cap (French); Kaapse Grasvogel (Dutch); Kap-Grassänger (German); Felosa do Cabo (Portuguese).
Recommended citation format: Tippett RM 2026. Cape Grassbird Sphenoeacus afer. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/07/08/cape-grassbird-sphenoeacus-afer/
List of bird species in this format is available here.
Cover image of Black-necked Grebe by Ryan Tippett – Carnarvon district, Northern Cape
Grebes are the sole members of the Family PODICIPEDAE. The family has a cosmopolitan distribution, occurring on all the world’s continents except Antarctica. Grebes are specialised for life on and under the water. Their legs are set so far back on the body that they have great difficulty walking on land. The toes are lobed for swimming. They also have sharp, pointed bills, short wings and tails, and soft, silky plumage. The family contains 22 species in 6 genera. 3 species have been recorded from southern Africa.
Identification
The Black-necked Grebe is an attractive species with distinct breeding and non-breeding plumage. The sexes are alike.
Black-necked Grebe Podiceps nigricollis Strandfontein Sewage Works, Western Cape Photo by Sybrand Venter
In breeding plumage, the forehead, crown, neck, mantle and scapulars are black. They have a patch of long, glossy golden-yellow feathers below and behind the eyes, extending past the ear coverts. The back is dark brown, and the rump is black. The short tail is black above and brownish below. The chin and throat are also black, while the upper breast is mottled with dark-chestnut and white. The sides of the breast and flanks are chestnut with black and grey mottling. The lower breast and belly are silky white, and the vent is pale brown. They have greyish-black bills and striking, bright red eyes. The legs and feet are blackish.
Black-necked Grebe Podiceps nigricollis Garden Route National Park, Western Cape Photo by Jean Hirons
In non-breeding plumage, they are greyish black above, and the ear coverts are grey, and the sides of the neck are black. The face, chin and upper throat are white. The lower throat is grey, and the remainder of the underparts are white. Their bills are also paler than in breeding plumage, and the feet are brown.
Juveniles resemble non-breeding adults, but are generally paler and browner. Additionally, the sides of the head are mottled, and the flanks are pale grey.
Black-necked Grebe Podiceps nigricollis Near Klerksdorp, North West Photo by Tony Archer
Black-necked Grebes in full breeding plumage are unlikely to be mistaken for another species. However, non-breeding birds are often confused with the Little Grebe (Tachybaptus ruficollis). Non-breeding Black-necked Grebes are much paler below than Little Grebes, and have longer and thinner necks. Little Grebes also have stubbier and less sharply pointed bills.
Status and Distribution
The Black-necked Grebe is uncommon to locally common in southern Africa. The species has a very wide distribution across much of Eurasia, North and South America, and Africa. In Africa, the Black-necked Grebe is found along the Mediterranean coast and at scattered sites south of the Sahel. In southern Africa, it mainly occurs in the central interior, from the highveld across to the drier south-western and western areas, with much of the population wintering along the Namibian coast.
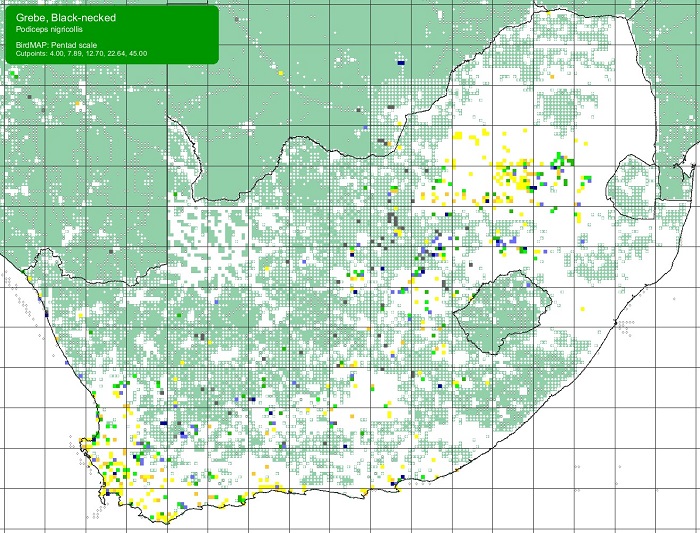
SABAP2 distribution map for Black-necked Grebe Podiceps nigricollis – December 2024. Details for map interpretation can be found here.
The Black-necked Grebe is not threatened. Its population has probably increased due to the widespread construction of man-made impoundments that offer ideal habitat. In southern Africa, the Black-necked Grebe now breeds in areas that formerly lacked suitable breeding habitat, such as the central Karoo.
Black-necked Grebe Podiceps nigricollis – Pair Carnarvon district, Northern Cape Photo by Ryan Tippett
Habitat
The Black-necked Grebe favours seasonal water bodies like large pans, ephemeral water bodies, and farm dams, particularly those that fill rapidly after heavy rainfall. They are less regular at more permanent waters. They are also frequently found on natural and commercial saltpans, and, in Namibia, in sheltered coastal bays. The Black-necked Grebe has a preference for open waters with sparse emergent vegetation, and they seldom enter areas with dense plant cover.
Ideal habitat at farm dam recently filled by rain. Carnarvon district, Northern Cape Photo by Ryan Tippett
Behaviour
Black-necked Grebes are usually seen in small groups or in pairs when breeding. Non-breeding birds may form large aggregations of 1000 or more, sometimes in association with Little Grebes. They roost on water at night.
Black-necked Grebe Podiceps nigricollis – A pair at the nest. Near Welkom, Free State Photo by Janet du Plooy
Black-necked Grebes are highly nomadic and are probably not resident anywhere in southern Africa. They breed sporadically when environmental conditions become suitable. Some of the population also move to inshore wintering areas along the Namibian coast. The only records at sea from the Northern Cape are from Port Nolloth. Black-necked Grebes are seldom recorded at sea off the Western Cape.
Black-necked Grebe Podiceps nigricollis Near Welkom, Free State Photo by Janet du Plooy
Foraging behaviour of the Black-necked Grebe in southern Africa is largely undescribed, except for observations of groups of 75 to 100 individuals feeding in association with Greater Flamingos. Dives for food, including small fish, tadpoles, aquatic insects and their larvae, as well as crustaceans and molluscs. Black-necked Grebes regularly swallow feathers, which are probably eaten to wrap fish bones for regurgitation in pellets.
Black-necked Grebe Podiceps nigricollis– Non-breeding birds at sea. Walvisbaai, Namibia Photo by Carel van der Merwe
The Black-necked Grebe breeds irregularly and opportunistically at flooded ephemeral pans, dams, and seasonal wetlands. In southern Africa, it has been recorded breeding from October to April, but the species may not breed at all for several years during drought conditions. The suitability of breeding sites in southern Africa is unpredictable. The Black-necked Grebe is monogamous and pairs may breed solitarily or in loose colonies. It seems that pair formation likely occurs only after a breeding site has been selected.
Black-necked Grebe Podiceps nigricollis – Nest Carnarvon district, Northern Cape Photo by Ryan Tippett
In Europe, Black-necked Grebes perform a range of ritualised and coordinated threat and courtship displays. In southern Africa, the only described early courtship display, involves the pair facing each other, shaking their heads from side to side, then lowering their heads, first to one side and then the other, after which they glide across the water side by side, with heads raised, showing their white breasts. Once a nest site has been selected, the pair constructs 1 or several nest platforms, followed by an additional courtship display where the pair rear up with quivering wings, followed by copulation.
The nest is a floating mound of algae, bulrush leaves, and other soft water plants, usually on open water. It is anchored on underwater or emergent growth or even a submerged termite mound, and the nest cup is often barely above the water level. Nests can be flimsy or rather substantial, measuring up to 200 mm in diameter and up to 500mm thick, and new material is added throughout the incubation process. The nest is cooperatively built in under a week by both sexes.
1 to 7 eggs are laid per clutch at intervals from 1 to several days. The eggs are symmetrically oval with a conical point at both ends. They are Yellowish-white but become brown-stained from the nest. The incubation period lasts up to 22 days and incubation duties are shared by both sexes. The eggs are sometimes covered with nesting material when the pair leave the nest. The newly hatched young are brownish-black overall, with black crowns and black-and-white striped faces. They also have bare pink to scarlet spots on their lores and crown. The chicks are highly precocial and leave the nest immediately after hatching. They are carried on the backs by both parents who also feed them.
Breeding success in southern Africa is low due mainly to the predation of eggs and fluctuating water levels. Additionally, many nests are also destroyed by strong winds.
Black-necked Grebe Podiceps nigricollis – Nest Carnarvon district, Northern Cape Photo by Ryan Tippett
Further Resources
Species text adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Carel van der Merwe, Janet du Plooy, Jean Hirons, Lappies Labuschagne, Ryan Tippett, Sybrand Venter, and Tony Archer is acknowledged.
Other common names: Blacknecked Grebe, Eared Grebe (Alt. English); Swartnekdobbertjie (Afrikaans); iDada (Zulu); Grèbe à cou noir (French); Schwarzhalstaucher (German); Mergulhão-de-pescoço-preto (Portuguese); Geoorde Fuut (Dutch).
A list of bird species in this format is available here.
Recommended citation format: Tippett RM. 2026. Black-necked GrebePodiceps nigricollis. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/06/29/black-necked-grebe-podiceps-nigricollis/
We were delighted to be back at ringing at Ouberg from 30 May and 4 June 2026! This is in the mountains about 20 km northeast of the Little Karoo town of Montagu. There is a map at the bottom of this blog.
Photo: Richard Gie
Our first supper together, in the big all-purpose meeting room at the De Kuilen Campsite at Ouberg Private Nature Reserve. This is a great venue!
The weather was not especially kind, and the total of 139 birds of 30 species handled was pretty good in the conditions. What was impressive was that 24% of the birds handled were retraps; 33 retraps of 139 birds. It is encouraging to know that within a few years we will start to be able to estimate survival rates for some of the more frequently retrapped species.
Photo: John Yeld
Joël and Achot are each taking a bird out of a mist net. What is not obvious is that they are on opposite sides of the net. This only becomes clear when you zoom in on Joël …
Photo: John Yeld
… this is why mist nets are effective at catching birds.
Photo: Sue Gie
We had a Blue Moon during the expedition. This was the second full moon in May, and this is moonrise on the evening of 31 May. The first full moon of the calendar month had been on 1 May, and the second was on 31 May. The last time there was a Blue Moon was in August 2023, so they don’t happen often!
Photo: Sue Gie
And here is moonset the following morning, 1 June. Astronomically, the moon is past full, and the actual full moon in June 2026 is on the last day of the month!
Photo: Sue Gie
The bright colours of the Bokmakierie against the black tee-shirt are impressive!
Photo: John Yeld
We handled a total of 10 Bar-throated Apalis. of which four were retraps. One, with ring AS53820, was ringed on 21 October 2022, and then retrapped in 2023 on 15 February and 11 May, and we trapped it on 30 May 2026. So it is at least 3.5 years old.
Photo: Sue Gie
With just the wing to go on, not everyone will instantly know this as a woodpecker. It is actually a …
Photo: Joël Daniel-Roux
… female Cardinal Woodpecker. She was a retrap, having been ringed on 17 February 2023.
Photo: John Yeld
The Cape Weaver has nine primaries, the main flight feathers of the wing. The outer primaries of the bird above have smooth and unworn tips. As you work inwards, the inner primaries are starting to show traces of wear. Moult of the primary feathers of the Cape Weaver takes place of a period of three months, starting on average in late November and finishing in late February. The outer primaries are thus about four months old, whereas the inner primaries have experienced a further three months of wear and tear.
Photo: John Yeld
Familiar Chat being ringed.
Photo: John Yeld
Familiar Chat. The light brown tips of the coverts, the feathers that partially cover the primaries and the secondaries have light brown tips. That pattern tells us this is a young bird, that fledged after the most recent breeding season, probably spring last year.
Photo: John Yeld
Familiar Chat. We take lots of photos of plumage, because when we retrap it, we can see how rapidly a young bird loses that the last of its juvenile plumage, and is indistinguishable from the adults.
Photo: John Yeld
Mist nets set up in the early evening in the hope of catching a nightjar.
Photo: John Yeld
Taking the nets down. No nightjar successes!
Photo: John Yeld
Ring-necked Dove. No primary feathers in moult.
Photo: John Yeld
Male Malachite Sunbird. Measuring the bill with dial callipers.
Photo: John Yeld
… and this is what he is concealing under his wing.
… and here is the Malachite Sunbird being released in slow motion
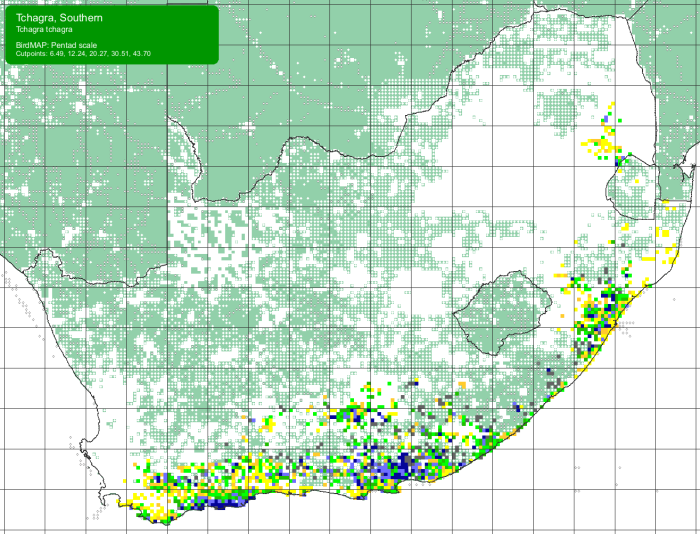
This is the first Southern Tchagra to be ringed at Ouberg, and only the third record here!
The SABAP2 map shows that Ouberg is close to the western edge of the usual range of the Southern Tchagra. Ouberg is almost due north of Cape Agulhas, the southern tip of Africa. In the map, the blue grid cells show the core of the range. In the yellow grid cells, it occurs irregularly. This is a shy species, and not easily spotted. This distribution map shows the entire range of this species, endemic to southern Africa
The tchagra is also not in moult. But that is what we expect. It’s winter!!
Photo: Ettienne Kotze
Rock Martin, huddling together, and keeping a watchful eye on proceedings.
Photo: Ettienne Kotze
Pretty much the middle of winter, but it is not too chilly to have a blether outside!
Photo: John Yeld
Sunset at Ouberg. This is a beautiful spot in its own right, independent of the birds!
Future BDI bird ringing opportunities are listed here.
The next expedition to Ouberg is from Saturday 11 July to Thursday 16 July 2026. You can attend for as many days as you are able. To book, or to ask questions, contact Sue Gie (bookings@ouberg.co.za).
There is a list of all expeditions with blog reports like this here. Note especially that there is a Ringers’ Symposium planned for November at Nuwejaars Wetlands SMA. The dates for ringing at 6 to 12 November, with the three days 8 to 10 November being the core period, with presentations, starting around 10h30, so we can do some ringing in the morning! Contact Sue Gie (sue@gie.co.za) for more details.
Here is the map that shows the route from the town of Montagu to Ouberg Private Nature Reserve.
Cover image of Sociable Weaver by Ryan Tippett – Near Kenhardt, Northern Cape
The Sociable Weaver is a member of the Family PLOCEIDAE (Weavers and Allies). Other members of Ploceidae include the Weavers, Bishops, Widowbirds, & Malimbes. Birds in this family are small passerines. These birds have short to medium, conical, pointed bills, adapted for seed-eating. They also have relatively strong legs and feet. Most are gregarious and are known for their intricately woven nests. They are distributed throughout sub-Saharan Africa, Madagascar, the southern edge of the arabian peninsula, and into tropical Asia. The family Ploceidae contains 123 species from 16 genera. Most species are found in Africa.
Identification
The Sociable Weaver is a conspicuous and characteristic bird of arid regions.
Sociable WeaverPhiletairus socius Hopetown district, Northern Cape Photo by Les Underhill
In adults, the body is mostly dull brown, save for black scalloping on the mantle, back and flanks. The forehead and crown are a slightly darker brown, and the cheeks and sides of the neck are plain buff. The folded wings are dull brown with variable buff edging on the feathers. The lores, the feathers around the base of the bill and the throat patch are black. The bill is dark horn-grey. The eyes are brown and the legs and feet are dark grey.
Sexes differ slightly in plumage colouration. Females closely resemble males but have a smaller black throat patch.
Sociable WeaverPhiletairus socius – Juvenile Carnarvon district, Northern Cape Photo by Ryan Tippett
Juveniles are similar to adult females, but lack the black feathers around the bill and on the throat. They also carry duller scalloping on the flanks.
The Sociable Weaver is not likely to be confused with any other bird species in southern Africa.
Sociable WeaverPhiletairus socius Near Marydale, Northern Cape Photo by Cobus Elstadt
Status and Distribution
Sociable Weavers are endemic to southern Africa where they are locally common. Their range is centred on the Northern Cape Province and southern Namibia where they are much more abundant than elsewhere.
They range from the Etosha area of northern Namibia south through southwestern Botswana, to the northeastern parts of the Northern Cape, central North West Province and the extreme northern Free State. Sociable Weavers are absent from most of the Namib Desert, the area immediately south of Etosha in Namibia, and from the Ghaap plateau in the Northern Cape.
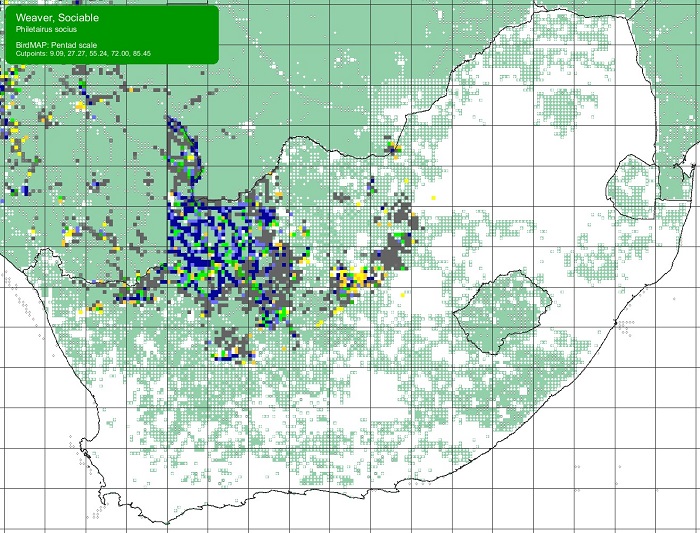
SABAP2 distribution map for Sociable WeaverPhiletairus socius – December 2024. Details for map interpretation can be found here.
The Sociable Weaver is not threatened. Its population has expanded substantially during the 20th century using telephone poles, electricity pylons, and other artificial structures as nest sites. The range could expand further if it were to accept alien trees for nesting. This may be happening in the Karoo, where they sometimes nest in larger Honey Mesquite (Prosopis glandulosa) trees. These trees are now a problematic invader across large swathes of the Northern Cape.
The range of the Sociable Weaver has contracted in parts of North West Province, the Free State, and the Ghaap Plateau in the Northern Cape, where woodland has been cleared for agriculture. In former times, the Sociable Weaver was hunted by the San people.
Habitat
The Sociable Weaver inhabits arid savanna and dry woodland, and is strongly associated with the Southern Kalahari vegetation type. It typically needs open savanna grassland with large trees for nesting. The Sociable Weaver is marginal in grassland, the Namib Desert and Mopane Colophospermum mopane woodland. Its spread into the largely treeless Nama Karoo and parts of the Namib have been permitted by artificial structures providing nesting sites. It is found in regions with a rainfall range of 80 to 600 mm per year.
Typical habitat in arid woodland dominated by thorn trees. Between Aus and Keetmanshoop, Namibia Photo by Les Underhill
Behaviour
The Sociable Weaver is highly gregarious at all times in colonies of about 6 to 300 birds. They are resident and sedentary, foraging birds seldom wander more than 1 or 2km from the nest. Each colony is associated with a communally built nest mass. The furthest recorded dispersal of young birds is 32.8 km, and there is very little movement of birds between colonies.
Sociable WeaverPhiletairus socius Sandveld Nature Reserve, Free State Photo by Ansie Dee Reis
They are often highly active at the nest, constantly flying up and down with building material or perching close by, calling almost constantly. They roost in the nest chambers, and 5 or more birds may roost in a single chamber, while other chambers remain empty. They return to the nest before sunset, perching in the canopy, lower branches of the nest tree, or in an adjacent tree. They fly in and out of several chambers before settling in one. Birds do not necessarily roost in the same chamber on consecutive nights unless breeding.
The underside of a Sociable WeaverPhiletairus socius nest showing the individual nest chambers. Carnarvon district, Northern Cape Photo by Ryan Tippett
Sociable Weavers are most well known for their huge nests, which are the World’s largest communal bird nests, and perhaps the most impressive structures built by any bird. The massive nest buffers birds from extreme temperatures. The air temperature within the nest is self-regulating. Day and night, through summer and winter, the nest temperature remains far more constant than the ambient air outside the nest. When it is cold outside, the inside temperature is around 20°C warmer, and the nest temperature increases with the number of occupants. Conversely, on hot summer days, the nest chambers can be as much as 20°C cooler than the air outside the nest.
Sociable WeaverPhiletairus socius Hopetown district, Northern Cape Photo by Les Underhill
Sociable Weavers drink regularly, flying up to 3.7 km from the nest to do so. They forage in flocks throughout the day, but mainly during the early morning and late afternoon. Groups forage on the ground, usually less than 2 km from the nest, sometimes joining feeding birds from other colonies. Birds on the ground move by quick hopping.
The diet of the Sociable Weaver is comprised mainly of seeds and arthropods, but they will also eat the soft lower nodes of grasses, as well as flowers and fruit. They mainly eat grass seeds, but legume seeds are also consumed. Arthropods are mostly eaten in summer, with termites forming the bulk, but caterpillars, moths, small grasshoppers, beetles, and ants are also taken.
Sociable WeaverPhiletairus socius Mokala National Park, Northern Cape Photo by John Todd
Sociable Weavers breed in any month, and they generally only breed in response to rain, with at least 20 mm of rain required to stimulate breeding. The breeding season may last up to 9 months in wet years with up to 4 broods per season. However, they may not breed at all during drought periods. Sociable Weavers are colonial, cooperative breeders, but are not genetically monogamous.
The nest is constructed by both sexes, but males are far more productive builders. The nest is an enormous, communal structure, sometimes measuring more than 7 m across, weighing more than 1 ton and containing over 250 nest chambers. Some larger nests are known to be over 100 years old. The entire structure consists of 2 well-defined parts: a dome-shaped superstructure (roof), usually built of fairly large, thorny twigs, long grasses and stems. It is positioned mostly above the supporting branch. Beneath it is a substructure of dry grasses, within which numerous separate nest chambers are embedded, each connected to the exterior by a vertical tunnel up to 250 mm long and 60-70 mm in diameter. Prior to egg-laying, the nest chamber is lined with soft, dry materials such as grass seed-heads, everlasting (Helichrysum spp.)leaves, and feathers. The Helichrysum leaves are strongly aromatic and are thought to guard against mites and other nest parasites. Nests are placed anywhere from 2 to 16 meters above the ground, and are usually north-facing.
A massive Sociable WeaverPhiletairus socius nest in a Camel Thorn Near Groblershoop, Northern Cape Photo by Ryan Tippett
The entire structure is maintained year-round. Building material is often collected from fallen portions below the nest, or brought back from foraging trips. Ideal nesting trees have a strong, more or less horizontal support branch with clear access from below. Nests are most often constructed in Camel Thorn Acacia erioloba trees and also in Grey Camel Thorn V. haematoxylon, Umbrella Thorn V. tortillis, Shepherds-tree Boscia albitrunca, Quiver-tree Aloe dichotoma, Karee Rhus lancea, Buffalo-thorn Ziziphus mucronata and Mopane Colophospermum mopane trees. Rarely in alien Eucalyptus spp. and mesquite (Prosopis spp.). Nests are also regularly built on telephone poles and, less often, on electricity and railway pylons, tank stands, wind-pumps, buildings, and natural rock faces.
Sociable WeaverPhiletairus socius Near Prieska, Northern Cape Photo by Ryan Tippett
2 to 6 (usually 3 or 4) eggs are laid per clutch at 1-day intervals. Intraspecific brood parasitism (egg dumping) is known to sometimes take place. Incubation usually starts with the first-laid and takes up to 14 days to complete. Incubation is done by both sexes, and sometimes by helpers. Adults and helpers roost in the nest chamber during both incubation and nestling periods, but the helpers often move to an adjacent chamber if too crowded. Hatching is asynchronous, however, the whole brood usually fledges on the same day or at least, within 2 days. The fledging period takes 21 to 24 days and the young are brooded continuously by both parents for their first 2 weeks. They are fed equally by both parents, and by up to 9 helpers of both sexes. Helpers are usually young from previous broods, but sometimes unrelated.
Nests of the Sociable WeaverPhiletairus socius are often placed in Quiver Trees . Carnarvon district, Northern Cape Photo by Ryan Tippett
Sociable Weaver colonies can be heavily impacted by predation. Snakes like the Cape Cobra Naja nivea, Boomslang Dispholidus typus, and Common Egg-eater Dasypeltis scabra can have a significant toll on breeding success. Many nestlings, fledglings, and adults are eaten by raptors, including Gabar Goshawk, Pale Chanting Goshawk, African Harrier- Hawk, Red-necked Falcon, and Pygmy Falcon.
Nest chambers are frequently usurped by mud-nesting wasps, nesting Pygmy Falcons, Red-headed Finches and Rosy-faced Lovebirds, and used as roosts by Ashy Tits, Familiar Chats, Acacia Pied Barbets, and Pearl-spotted Owlets.
Sociable WeaverPhiletairus socius Fish River Canyon National Park, Namibia Photo by Attie van Aarde
Further Resources
This species text is adapted from the first Southern African Bird Atlas Project (SABAP1), 1997.
The use of photographs by Ansie Dee Reis, Attie van Aarde, Cobus Elstadt, John Todd, Les Underhill, and Ryan Tippett is acknowledged.
Other names: Versamelvoël (Afrikaans); Républicain social (French); Republikeinwever (Dutch); Siedelweber (German); Tecelão-sociável (Portuguese).
Recommended citation format: Tippett RM. 2026. Sociable Weaver Philetairus socius. Biodiversity and Development Institute. Available online at https://thebdi.org/2026/06/19/sociable-weaver-philetairus-socius/
List of bird species in this format is available here.