View the above photo record (by Tyrone Ping) in FrogMAPhere.
Find the Natal Cascade Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Heleophrynidae
NATAL CASCADE FROG – Hadromophryne natalensis
Hewitt, 1913
Habitat
H. natalensis inhabits clear, swift-flowing streams in mountainous terrain; these waters flow through wooded and forested habitat and have headwaters in montane grassland. Annual rainfall in these habitats is 800–2700 mm. Adults often frequent waterfalls and cascades, where they may be found beneath submerged rocks, in rock cracks, in caves, or sometimes in exposed positions on wet rock faces. Tadpoles live on rocky substrates in swift-flowing streams; when disturbed they take cover beneath rocks or in cracks (Boycott in prep.).
This species occupies both Forest and Grassland biomes. Vegetation types include Afromontane Forest, Wet Cold Highveld Grassland, Moist Upland Grassland, North-eastern Mountain Grassland, Afro Mountain Grassland and Short Mistbelt Grassland.
Behaviour
Young frogs have been collected under stones and on moss covered boulders in shallow headwater streams in May and August in the Woodbush Forest and the Wonderwoud. Three adult female specimens were found in a wet rocky recess at the base of a waterfall in October in Malolotja Nature Reserve, Swaziland. These observations indicate that at least some individuals in the population remain in the vicinity of the breeding habitat after the presumed breeding period has passed. In the KwaZulu-Natal Drakensberg, adults were found in a soil pit and in a hole in a road cutting in grassland, some distance from the nearest streams and forests (Bourquin and Channing 1980; Channing 2001).
Breeding usually takes place in late summer (March–May) when stream flow is reduced, and before winter temperatures become severe (pers. obs.). Males usually call from hidden sites beneath clusters of boulders in stream beds, under the roots of large trees growing in streams, amongst boulders that have created a cascade of rushing water, in rock cracks and crevices in waterfalls, and on cliff faces and rocks located close to waterfalls and cascades. The species has not been heard calling while submerged and males do not call in close proximity to one another, even during peak breeding periods (Boycott in prep.).
The eggs and oviposition sites of H. natalensis have not been described, but it is unlikely that its breeding biology differs greatly from that of the related Heleophryne species.
The forested ravines and high altitude montane grasslands, which are the natural habitat of H. natalensis, are mostly protected within remote wilderness areas. However, afforestation with exotic trees, particularly in Mpumalanga and Limpopo provinces, has substantially reduced the non-breeding habitat and caused perennial streams to dry up, effectively eliminating entire breeding populations in the process (L.R. Minter pers. obs.). Additional threats to certain isolated populations comprise the damming of mountain rivers as has occurred in the Lesotho Highlands Project, water extraction and the introduction of alien fish species into the habitat. Nonetheless, owing to the wide distribution of this species, it is not considered to be a conservation priority. It occurs in several private and public protected areas, including Ukahlamba-Drakensberg Park, a World Heritage Site.
H. natalensis is endemic to the atlas region. It occurs throughout the Drakensberg and Maluti mountains and along the great escarpment of South Africa, Swaziland and Lesotho. The recorded altitudinal range is 580–2675 m (Boycott in prep.).
The atlas records are reliable but incomplete, especially for most of Lesotho. This species should be easy to detect since the tadpoles are easily identified and are present in streams throughout the year. Channing (2001) expressed the opinion that this taxon contains a number of cryptic species and that field and laboratory investigations should be undertaken to explore this possibility.
Distribution of Hadromophryne natalensis. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Natal Ghost Frog (Alternative English Name); Natalse Snelstroompadda (Afrikaans)
Recommended citation format for this species text:
Boycott RC, Tippett RM. Natal Cascade Frog Hadromophryne natalensis. BDI, Cape Town. Available online at http://thebdi.org/2021/12/20/natal-cascade-frog-hadromophryne-natalensis/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Boycott RC 2004 Hadromophryne natalensis Natal Cascade Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Find the Rose’s MountainToadlet in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
Identification
Rose’s Mountain Toadlet is a small Bufonid with an elongated body that reaches a length of c.28 mm in males and 39 mm in females (Channing 2001). The dorsum is smooth and soft with scattered blister-like ridges and warts on the back and sides, while the posterior region of the ventrum has heavily granulated skin (Passmore and Carruthers 1995). The pupil is horizontally elliptical. Interestingly, this species lacks external or middle-ear structures (Grandison 1980) and apparently has no advertisement call. The parotid glands are distinct and have an inverted pear shape (when viewed from the rear). There is no webbing between the toes, and the hind limbs are relatively short and better adapted for walking and running than for jumping (Poynton 1964; Wager 1965).
The upper body surface is variably shaded in grey to brown, interspersed with darker spots and blotches. A light vertebral stripe or band and a pair of light dorsolateral bands are usually present. The parotid glands are generally orange to red and markings of the same colour may be present elsewhere on the upper body surface. The underside is white with variable grey shading and some darker markings may be present. These frogs often take on a uniformly blackish appearance when in dark surroundings.
Rose’s Mountain Toadlet most resembles C. magistratus and C. selenophos as these species also lack a tympanum and external or middle-ear structures. Two other Capensibufo species, namely, C. deceptus and C. tradouwi are also similar, but they posess a tympanum and produce advertisement calls (See ‘Similar Species’ below).
Rose’s Mountain Toadlet Capensibufo rosei Silvermine, Western Cape Photo by Luke Kemp
Habitat
Rose’s Mountain Toadlet is restricted to mountains where it occurs in undisturbed Mountain Fynbos. Although it is usually associated with the generally flatter topography on mountain tops, C. rosei has also been recorded from low-lying plateaus on the southern Cape Peninsula.
Breeding takes place in small, shallow, temporary pools of standing water that form during the winter rainy season. The pools can vary in size from c.20 × 20 × 2 cm deep to 250 × 150 × 6 cm deep. The vegetation surrounding breeding pools is usually dominated by restios. Pool substrates generally consist of a dark humic (peaty) or mossy layer on a sandy or rocky base and vary from light grey to almost black depending on the humic content. A breeding site can consist of one or more small pools in a seepage area of, for example, 100 m2. The species is not known to breed in pools of moving water associated with mountain streams.
Behaviour
Rose’s Mountain Toadlet has been seen in all four seasons of the year, but breeds only in late winter and spring. The commencement of breeding is dependent on rainfall and the formation of small, shallow pools of water. There are relatively few published records of C. rosei breeding activity. Spawn has been found in late July to early October.
An unusual characteristic of Capensibufo rosei is the absence of middle-ear elements (Grandison 1980), and it along with C. magistratus and C. selenophos appear to be the only southern African amphibians that lack a voice. Large, dense breeding aggregations and repeated use of the same site may facilitate meeting of the sexes. For example, at least 150–200 adults (and spawn) were observed in a partially divided breeding pool with overall dimensions of 250 × 150 cm and up to 6 cm deep (pers. obs.). (A similar observation was made by Power and Rose, who also described the eggs, tadpoles and metamorphosis of C. rosei (Rose 1962). During the breeding season, a bright pink, oval patch appears below the vent in both sexes (Power and Rose 1929), and a distinct swelling develops in the cloacal region in breeding males. The functions of these structures are not known.
These toads appear to be particular in their choice of breeding site. They have been found to use the same site on a few occasions during a breeding season, despite the presence of other similar small pools in the general vicinity (pers. obs.; southern Cape Peninsula).
The large, pigmented eggs are laid in a single transparent jelly tube that resembles a string of beads in that there is a distinct narrowing of the tube between the egg capsules (Power and Rose 1929). The clutch size is reported to be <100 eggs (Grandison 1980) but the typically dense breeding aggregations can lead to the deposition of thousands of eggs in a single pool. Adults in the breeding pool sometimes hide under the egg masses when disturbed.
The embryos take up to 12 days to leave the egg capsules. The tadpoles are dark in colour and attain a length of about 21 mm, of which more than half comprises the tail. They are benthic in habit, and have been observed feeding on egg capsules, decaying eggs and dead adults. Metamorphosis takes about six weeks, depending on temperature and the availability of food and water. The tiny toads are about 6.25 mm in length when they leave the water.
The predators and prey of this species have not been recorded.
Rose’s Mountain Toadlet Capensibufo rosei Silvermine, Western Cape Photo by Felicity Grundlingh
Status and Conservation
Status
C. rosei was previously listed as Restricted (Branch 1988) and is presently listed as Vulnerable (Harrison et al. 2001; this publication). It is legally protected by Nature Conservation Ordinance 19 of 1974, but is not listed by CITES.
Rose’s Mountain Toadlethas a restricted and fragmented distribution comprising about six subpopulations (sensu IUCN criteria). While most C. rosei habitat is situated in pristine mountain areas, habitat degradation has led to <20% habitat loss over the last 30 years. However, in some areas, the quality of C. rosei habitat is likely to have improved with systematic alien vegetation control programmes. For example, the previously degraded habitat of C. rosei in the Silvermine area on the Cape Peninsula (Boycott 1988e) now falls within Cape Peninsula National Park, and the pine plantations and dense stands of alien vegetation have been removed.
C. rosei is known to occur in the following protected areas: Cape Peninsula National Park (including the former Silvermine Nature Reserve and Cape of Good Hope Nature Reserve), Hottentots Holland, Kogelberg, Fernkloof, Maanschynkop, Vogelgat, Limietberg and Riviersonderend nature reserves.
Threats
In some places, one or more of the following factors have led to degradation or loss of habitat, or are a potential threat to populations of C. rosei. These include invasive alien vegetation, plantations, wildflower farming, alteration of drainage patterns, too frequent fires, building developments such as roads and dams, and urban development on the southern Cape Peninsula. The most pressing of these threats is probably the spread of invasive alien vegetation that eliminates fynbos vegetation and reduces water resources.
The tendency of C. rosei to form dense breeding aggregations at specific sites increases its vulnerability, since the destruction or degradation of a breeding site seriously threatens the continued existence of that local population.
Amphibian declines in some parts of the world have taken place in pristine, high-altitude areas, and this has led to speculation supported by some scientific evidence that global environmental factors are responsible (Beebee 1997). In particular, climatic and atmospheric changes brought about by widespread atmospheric pollution, increased ultraviolet radiation and global warming appear to be the prime causes. These factors may also lower amphibians’ resistance to disease. There is no evidence, at present, that global factors have caused declines in southern African amphibian populations, but such threats may be significant.
Recommended conservation actions
The distribution and conservation status of C. rosei is monitored by the Western Cape Nature Conservation Board (De Villiers 1997a) as part of a threatened species monitoring programme. This helps ensure that appropriate recommendations can be made to promote the conservation and wise management of this species and its habitat. Management of C. rosei habitat is included in the management plans of the respective protected areas and mainly concerns the control of invasive alien vegetation.
Research is underway to investigate the genetic relationships between the various geographically fragmented subpopulations of C. rosei. Life history studies of this species are also in progress. A captive breeding programme for C. rosei is not recommended.
Distribution
Rose’s Mountain Toadlet is endemic to the winter-rainfall region of the Western Cape Province. It has a limited and patchy distribution. Its populations are restricted to the Cape Peninsula and mountains southwest of the Breede River valley, which extends from the Tulbagh/Worcester valley south-eastward to Witsand on the south coast, and the Nuwekloof gap that separates the Bain’s Kloof Mountains from the mountains to the north (Grandison 1980). Recorded localities vary in altitude from 60 m on the southern Cape Peninsula to 1600 m, although >80% of localities are above 400 m. The areas in which this species occurs receive 600–>3000 mm of rain per annum.
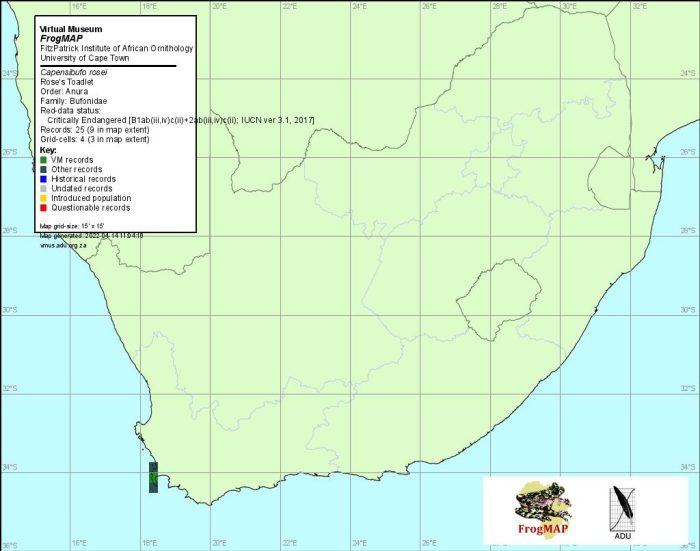
Distribution of Capensibufo rosei. Taken from the FrogMap database, April 2022.
The Rose’s Mountain Toadlet was discovered on the Cape Peninsula (Hewitt 1926a), where it is known from the following localities: Table Mountain (3318CD), Muizenberg Mountain (type locality), Kalk Bay Mountain, Silvermine Valley, Chapman’s Peak and Kommetjie area (all in 3418AB), and Cape of Good Hope area (3418AD). However, the main part of its distribution range lies elsewhere, and includes the following localities: Bailey’s Peak in Limietberge Mountains (a.k.a. Bain’s Kloof Mountains) and Hawekwa Mountains (3319CA), Riviersonderend Mountains (3419BB) including Jonaskop (3319CD) and Kanonberg (3419BA), Klein River Mountains (3419AD), Jakkals River catchment area, Nuweberg Mountain near Viljoen’s Pass, Groenlandberge Mountains, Landdroskop in the Hottentots Holland Mountains (all in 3419AA), Elgin Forest Reserve, and Kogelberg State Forest (3418BB).
Capensibufo rosei has a geographically fragmented distribution pattern that, based on current knowledge, appears to comprise six populations: Table Mountain, southern Cape Peninsula, Hottentots Holland/Kogelberg mountain complex, Dutoitsberge/Limietberge mountain complex, Riviersonderend Mountains, and Klein River Mountains.
In summary, C. rosei has been found in a total of 10 quarter-degree grid cells, all of which were recorded for the first time before 1990. However, since 1996 the species has been recorded in only five of these cells: 3418AB, 3418AD, 3319CA, 3419AA and 3419AD. Most of the latter records were obtained in 2000 during a threatened frog monitoring programme (De Villiers 1997a).
This small, cryptic toad is difficult to find in its remote montane habitat. However, the distribution data are accurate and it is unlikely that the limits of the species’ range will be extended by further surveys.
Further Resources
The use of photographs by Felicity Grundlingh, Luke Kemp, and Megan Loftie Eaton is acknowledged.
Other Common Names: Gestreepte Skurwepadda (Afrikaans)
Recommended citation format: de Villiers, AL; Tippett, RM. (2025). Rose’s Mountain ToadletCapensibufo rosei. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/20/roses-mountain-toadlet-capensibufo-rosei/
This species text has been updated and expanded from the text in the 2004 frog atlas: de Villiers, AL. (2004). Rose’s Mountain Toadlet Capensibufo rosei. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Find the Tradouw MountainToadlet in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
Identification
The Tradouw Mountain Toadlet is a small Bufonid that attains a maximum size of 48mm. It has a toad-like appearance, but unlike true toads, it has smooth skin with blister-like ridges and warts, and the body is somewhat elongated.
The dorsal surface colouration varies from dark reddish-brown to black, with shades of grey and with scattered dark blotches. A pale vertebral stripe is often, but not always, present. The parotid glands are prominent and orange to red in colour. A row of orange-red warts is found along the flanks. The tympanum is visible and is smaller than the eye. The underside is white with variable dark grey mottling.
Tradouw MountainToadletCapensibufo tradouwi Wolfberg Arch, Western Cape Photo by Paul Bester
Habitat
The Tradouw Mountain Toadlet occurs at high altitudes in mountainous terrain where it inhabits Mountain Fynbos and Grassy Fynbos. Most of its range lies within the western winter-rainfall region, but in the east extends into the zone of transition to summer rainfall.
Breeding takes place on mountain slopes and valley bottoms in shallow pools in seepage areas, moist depressions, vleis and slow streams. A breeding site on the slopes of Matroosberg in the Hex River Mountains consisted of rock pools varying in depth from a few to several centimetres, in a large expanse of exposed bedrock above a waterfall. Clumps of reeds and grass growing in the cracks provided limited shade to the pools that, in most cases, were on the bedrock and exposed to direct sunlight (pers obs).
Behaviour
During the day, adults shelter under rocks but may also be found in the open near seepage areas. Diurnal activity is especially common during the breeding season, June–September, when males are often found walking about at the breeding site. A number of tadpoles, found in rock pools at Matroosberg toward the end of October 1982, had hindlimbs, indicating that spawning had taken place in September, assuming that C. tradouwi has a developmental period similar to C. rosei, that is, 25–30 days (pers. obs.).
At the end of October, Tradouw MountainToadlets were common at the Matroosberg locality, with >20 specimens seen over a two-day period. They were found under stones lying on bedrock in seepage areas, and even on the high, dry slopes. A somewhat later breeding period was recorded at the eastern-most locality in the Tsitsikamma Mountains. Here, males were observed calling in late November 1993, in close proximity to small pools containing tadpoles (M. Burger pers. comm.).
Tradouw MountainToadletCapensibufo tradouwi Op de Berg, Western Cape Photo by Nick Helme
The males call from the edge of shallow pools, often from concealed positions (Channing 2001). Amplexus is axillary. The eggs are unusual in that they are laid singly; in most bufonids, including C. rosei, they are laid in strings. The clutch size is c.60 eggs which are 2 mm in diameter and black at one pole (Channing 2001).
The tadpoles are dark, with unusually long undulating tails, and are benthic in habit (Passmore and Carruthers 1995). At the Matroosberg site, small black tadpoles were densely packed in the pools in plain sight. In some of the larger pools in the bedrock, as many as 100 tadpoles were found, while in the smallest pools, several tadpoles were present. Tadpoles were also found sheltering under small rocks that were lying in some of the pools.
Status and Conservation
The Tradouw MountainToadlet has a wide distribution in the southwestern and southern parts of the Western Cape Province, in remote mountainous areas. It occurs in a number of protected areas, including Cederberg Wilderness Area, Grootvadersbosch Nature Reserve and Formosa Conservation Area, and does not appear to be threatened. Nevertheless, there have been few recent observations of C. tradouwi, and it is recommended that surveys be undertaken to confirm its current status.
Distribution
The Tradouw MountainToadlet is endemic to the Cape fold mountains in the southwest and south of the Western Cape Province (Grandison 1980), with the most easterly locality marginally entering Eastern Cape Province at Misgund (3323CD) in the Tsitsikamma Mountains. The species occurs north and east of the Berg and Breede river valleys, from the Matsikammaberg (3118DD) near Vanrhynsdorp in the north, southward to the Hex River Mountains, and eastward along the Langeberg mountain range to the Outeniqua, Swartberg and Kammanassie ranges. The species is recorded at altitudes of 1000–1600 m.
The atlas data are accurate but incomplete.
Distribution of Capensibufo tradouwi. Taken from the FrogMap database, April 2022.
Further Resources
The use of photographs by Nick Helme and Paul Bester is acknowledged.
Other Common Names: Tradouw-bergskurwepadda (Afrikaans)
Recommended citation format: Boycott, RC; Tippett, RM. (2025). Tradouw MountainToadletCapensibufo tradouwi. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/20/tradouw-mountain-toadlet-capensibufo-tradouwi/
This species text has been updated and expanded from the text in the 2004 frog atlas: Boycott, RC. (2004). Tradouw MountainToadletCapensibufo tradouwi. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
View the above photo record (by Gregg Darling) in FrogMAPhere.
Find the Eastern Leopard Toad in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
EASTERN LEOPARD TOAD – Sclerophrys pardalis
(Hewitt, 1935)
Habitat
S. pardalis inhabits grassy or open bushveld areas, parks and gardens in the Thicket, Grassland and Savanna biomes, and uses large, permanent water bodies for breeding (Lambiris 1989a). It appears to prefer ecotonal habitat with moist cover. Records from Stutterheim and Woody Cape are close to patches of Afromontane Forest and Coastal Forest, respectively. The south coast populations may occur, to a limited extent, in fynbos vegetation types.
Behaviour
Like its western counterpart, S. pardalis is a late-winter or spring breeder. Choruses are typically heard from August to September, but calling is also recorded from November through to January (Burger 1997). Channing (2001) also noted calling in November. Calling behaviour was studied by Passmore (1977b) who confirmed earlier reports that this large toad usually calls while floating in the water in a manner similar to Schismaderma carens. Lambiris (1989a) noted that in Grahamstown, males call in water deeper than 80 cm while grasping emergent vegetation with one hand.
This is the largest southern African bufonid attaining a snout–vent length of 147 mm and a mass of 410.7 g (Branch 1995). Captive specimens feed readily on crickets, grasshoppers, small mice (pers. obs.) and lizards (Channing 2001). It is presumed that they feed on a variety of arthropods in the wild.
Status and Conservation
This species has been recorded from Addo Elephant National Park (Branch and Braack 1987), Groendal Wilderness Area, Thomas Baines Nature Reserve and Woody Cape Nature Reserve, and is likely to occur in several other protected areas. S. pardalis is assigned to the category Least Concern (Harrison et al. 2001; this publication).
Although it may still be common at some localities, the impact of agricultural and urban development has led to fragmentation and degradation of this species’ habitat, a reduction in population numbers and possibly even local extinctions. During the breeding season, large numbers are killed by motor vehicles on roads situated near wetlands: for example, 113 S. pardalis were killed in one night along an 8-km stretch of road near Port Elizabeth (Branch 1980). The conservation actions recommended for S. pantherinus would also be appropriate for S. pardalis, especially in built-up areas.
Until recently, S. pardalis was regarded as having a disjunct distribution, with one population restricted to the southwestern coastal areas of Western Cape Province and the other confined, mainly, to Eastern Cape Province (see map in Passmore and Carruthers 1995). However, the western population was raised to the status of a full species, Sclerophrys pantherinus, by Poynton and Lambiris (1998) on the basis of morphological differences, a decision supported by Eick et al. (2001; see S. pantherinus species account for further discussion).
S. pardalis is nearly endemic to the Eastern Cape Province with only a few records in the Western Cape and KwaZulu-Natal. The isolated record from Albert Falls (2930AD) in KwaZulu-Natal (Lambiris 1989a, 1994) requires confirmation by acoustic or genetic studies.
This toad is most common along the coast from Port Elizabeth northwards to East London. Inland records include Grahamstown, Kei Road (the type locality) and Stutterheim. Branch (1990) also recorded it from the Amatola Mountains and Katberg area. A recent record from Umtata (3128DB) represents another interesting, apparently isolated, population. Similarly, the recent Karatara (3322DD) and Knysna (3423AA) records are interesting in that they confirms the southwestern limit of the species’ range in the vicinity of Wilderness (Branch and Braack 1987). The atlas data are accurate but incomplete.
Distribution of Sclerophrys pardalis. Taken from the FrogMAP database, April 2022
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Oostelike LuiperdSkurwepadda (Afrikaans)
Recommended citation format for this species text:
Burger M, Tippett RM. Eastern Leopard Toad Sclerophrys pardalis. BDI, Cape Town. Available online at http://thebdi.org/2021/12/20/eastern-leopard-toad-sclerophrys-pardalis/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Burger M 2004 Sclerophrys pardalis Eastern Leopard Toad. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Christopher Willis) in FrogMAPhere.
Find the Flat-backed Toad in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
FLAT-BACKED TOAD – Sclerophrys pusilla
Hallowell, 1854
Habitat
In the atlas region, this species inhabits various vegetation types within the Savanna and Grassland biomes, on the Zululand coastal plain (Lambiris 1989a), in the Swaziland lowveld and middleveld (Boycott 1992), and in the lowveld and bushveld of northeastern South Africa (Jacobsen 1989). It is usually associated with riverine habitats: in Swaziland the species is almost always associated with medium and large rivers and is seldom found at pans, borrow pits or dams. In Ivory Coast, the species occurs in both forest and savanna habitats.
Breeding habitat is riverine. During the dry season in Ivory Coast, the species breeds alongside rivers in small, shallow inlets and puddles created by rising and falling water, as well as in rock-pools, while in the wet season it moves away from rivers to forage in gallery forest or forms new choruses in savanna, usually in periodic brooks (Rödel 2000).
At Pafuri, in the north of Kruger National Park, adults and juveniles have been found sheltering under rocks, logs, piles of dead leaves and branches, and building debris (H. Braack pers. comm.).
Behaviour
Breeding takes place from September to January in Kruger National Park (H. Braack pers. comm.), while in Ivory Coast it occurs throughout the year (Rödel 2000). Some authors recorded an increase in breeding activity during dry periods (Channing 1989; Rödel 2000).
In Swaziland, and elsewhere in the atlas region, the species calls from well-concealed positions amongst reeds, grass or rocks, next to or in rivers and streams where there is fast-flowing water (Jacobsen 1989; pers obs.). In Zimbabwe, calling from exposed sites has been observed (Lambiris 1989b). Calling takes place at night and during the day.
The calls are often antiphonal, with individuals calling at slightly different pitches so that alternate calls are readily distinguishable (Pienaar et al. 1976; Lambiris 1989a). Females approach calling males and make contact prior to amplexus (Channing 2001). Amplexus is axillary. Wager (1986) recorded eggs 1.2 mm in diameter in a single jelly tube 3 mm thick, while Tandy and Keith (1972) asserted that all African Sclerophrys species produce two egg strings. In Ivory Coast, Sclerophrys pusilla produces two egg strings simultaneously from the cloaca, containing 2000–8000 eggs, 1.5 mm in diameter (Rödel 2000). The egg strings are deposited at the bottom of a pool or wound around stones or twigs. Tadpoles reach metamorphosis within six weeks in captivity and in as little as two weeks in the wild (Rödel 2000).
Sandflies are known to feed on the blood of Sclerophrys pusilla (Braack et al. 1981), while juveniles and adults are preyed on by the Common Night Adder Caususrhombeatus (Pienaar et al. 1976). In Ivory Coast, the Spotted Night Adder Causus maculatus and a scorpion Pandinus imperator were recorded as predators (Rödel 2000). In savanna pools in Ivory Coast, tadpoles of the Crowned Bullfrog Hoplobatrachus occipitalis feed on the tadpoles of Sclerophrys pusilla (Rödel 2000), while in the atlas region they are sometimes taken by Mozambique Tilapia Oreochromis mossambicus (H. Braack pers. comm.). Adults feed mainly on ants, but also on beetles and alate termites (Rödel 2000).
Sclerophrys pusilla is widespread and common, occurring in several national parks and other protected areas in South Africa and Swaziland (Jacobsen 1989; Lambiris 1989a; Boycott 1992). Much of its habitat is well protected in cattle and game farming areas; consequently the species is not threatened.
Sclerophrys pusilla is widespread, occurring in East Africa and southern Africa. The species is found in northeastern South Africa, reaching its southern limits in Swaziland and northern KwaZulu-Natal. In Swaziland it occurs at altitudes of 90–790 m, and in South Africa, from 40–1700 m (Jacobsen 1989). These toads occur quite far inland in Swaziland and adjacent South Africa, where their distribution range extends up the larger river valleys, including the Nkomati, Mlumati, Elands, Olifants and Limpopo rivers.
This species is easily identified by its call. The atlas data are accurate but incomplete.
Distribution of Sclerophrys pusilla. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gestreepte Skurwepadda (Afrikaans)
Recommended citation format for this species text:
Boycott RC, Tippett RM. Flat-backed Toad Sclerophrys pusilla. BDI, Cape Town. Available online at http://thebdi.org/2021/12/18/flat-backed-toad-sclerophrys-pusilla/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Boycott RC 2004 Sclerophrys pusilla Flat-backed Toad. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Find the Namaqua Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicipitidae
Identification
The Namaqua Rain Frog female reaches a length of 45mm; males are smaller. The dorsal surface is dark brown with lighter sides, and the underside is white and smooth. The eyes are relatively large. Namaqua Rain Frogs have well-developed double tubercles beneath the toes.
Namaqua Rain FrogBreviceps namaquensis West Coast National Park, Western Cape Photo by Trevor Hardaker
Habitat
The Namaqua Rain Frog appears to favour deep, scrub-covered red sands in the Succulent Karoo Biome. Specifically in the Strandveld and Lowland Succulent Karoo vegetation types. The harsh, brackish soils of the Knersvlakte are avoided. Unlike B. macrops, this species is not confined to coastal dunes and substantial populations were encountered in the interior.
Behaviour
Little is known about the life history of the Namaqua Rain Frog. It is presumably similar to that of other members of the genus.
Namaqua Rain FrogBreviceps namaquensis North of Papendorp, Western Cape Photo by Erlo Brown
Calling takes place mainly in winter and spring, during and following good rain. Sporadic calling has also been noted in early summer and autumn in the western Richtersveld and during periods of heavy mist following recent rain (pers. obs.). Males call from concealed positions beneath vegetation, in scrub-covered sandy areas (pers. obs.; Channing 2001).
Advertisement calls of B. namaquensis, recorded by A. de Villiers at Rocherpan Nature Reserve, had an emphasized frequency of 1600 Hz rising to 2000 Hz. The calls were 160–165 ms in duration and were repeated at a rate of about one call per second (L.R. Minter pers. comm.). The calls of this species are higher pitched and much shorter than the calls of B. macrops (see species account). It makes a squeaky distress call; listen to it here.
Status and Conservation
The Namaqua Rain Frog is protected by ordinances of the Northern Cape and Western Cape provincial nature conservation agencies. The species occurs in the Corridor West (2917AA, CA) area of the Richtersveld National Park and in the Rocher Pan Nature Reserve (A. de Villiers pers. comm.).
In the southern half of its range, certain parts of its habitat have been exploited for agriculture and development, but in general this species is not under threat.
Namaqua Rain FrogBreviceps namaquensis Soutkloof farm, Western Cape Photo by Paul Bester
Because so little is known about the Namaqua Rain Frog, it is recommended that further studies on its distribution and general biology be undertaken. Regular monitoring of protected populations is advisable and would certainly yield new and useful information.
Distribution
The Namaqua Rain Frog is endemic to the atlas region. It occurs in the predominantly winter-rainfall Namaqualand region of the Northern Cape and Western Cape provinces, from near the Gariep (Orange) River in the north (2816DA) to the Melkbos district (3318CB, DA) in the south. V.C. Carruthers (unpubl. data) recorded a specimen of B.namaquensis disgorged by a snake near Elands Bay (3218AD). The escarpment and the Namaqualand and Richtersveld mountains form the eastern limit of this species’ distribution.
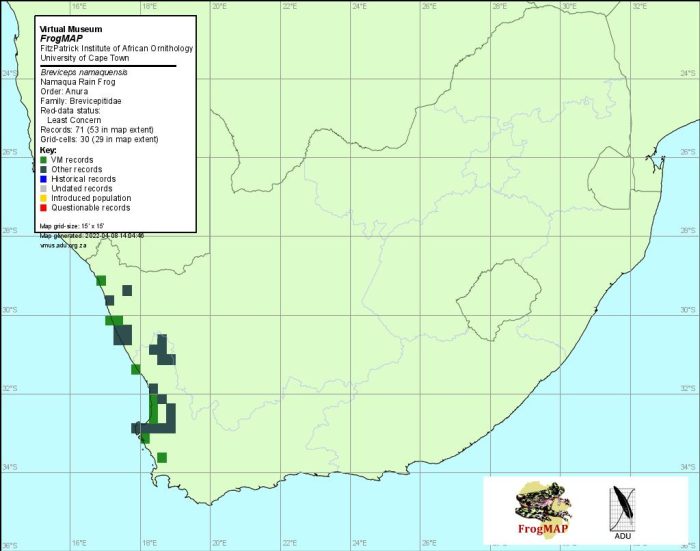
Distribution of Breviceps namaquensis. Taken from the FrogMAP database, April 2022.
In the northern part of its range in Port Nolloth (2917AC), Holgat River (2816DD), Alexander Bay (2816CB) and surrounding districts, B.namaquensis occurs in sympatry with B. macrops. It is sympatric with B. rosei in the southern reaches of its range. Morphological and call differences between the three species easily separate them, and the atlas data are reliable. Gaps in the recorded distribution of this frog probably indicate inadequate surveying rather than a real absence of the species.
Further Resources
The use of photographs by Erlo Brown, Paul Bester, Trevor Hardaker, and Vaughan Jessnitz is acknowledged.
Recommended citation format: Braack, HH; Tippett, RM. (2025). Namaqua Rain Frog Breviceps namaquensis. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/17/namaqua-rain-frog-breviceps-namaquensis/
This species text has been updated and expanded from the text in the 2004 frog atlas: Braack, HH. (2004). Namaqua Rain Frog Breviceps namaquensis. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
Find the Mozambique Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
The Mozambique Rain Frog attains a maximum length of 52mm. Females are larger than males. The body is rotund with short legs and small eyes.
In the atlas region, the Mozambique Rain Frog exhibits considerable intra- and inter-population variation in colour and markings. Most individuals have light paravertebral and dorsolateral patches (see illustrations in Passmore and Carruthers 1995) differing, in this respect, from the northern Mozambique populations of the species. Due to the presence of similar markings in B.sopranus, B. bagginsi and B. adspersus, this character is of little diagnostic value. This weakens the assertion, based mainly on the presence, absence or extent of dorsal markings (Poynton 1964; Poynton and Broadley 1985a; Lambiris 1989a), that mossambicus and adspersus hybridise extensively in sympatry. In addition, a study of advertisement call structure found little evidence of hybridisation between these species (Minter 1998). At present, these species are morphologically indistinguishable, and identification should be based on advertisement call structure.
Mozambique Rain Frog Breviceps mossambicus Marakele National Park, Limpopo Photo by A. Coetzer
Habitat
The Mozambique Rain Frog inhabits parts of the Savanna and Grassland biomes where the annual rainfall exceeds 700 mm per year. Coastal populations of B. mossambicus in KwaZulu-Natal and the Eastern Cape inhabit well-drained, sandy soils in Coastal Bushveld-Grassland, Coast-Hinterland Bushveld and Natal Lowveld Bushveld. At higher altitudes, this species occurs along the crest and slopes of the escarpment; vegetation types include Moist Sandy Highveld Grassland, Moist Upland Grassland, Short Mistbelt Grassland, and North-eastern Mountain Grassland. The soil is usually shallow, well drained, humus-rich, and often rocky.
Habitat in Coastal Grassland on well-drained, sandy soil. Western Shores, iSimangaliso Wetland Park, KwaZulu-Natal. Photo by Ryan Tippett
Behaviour
The Mozambique Rain Frog breeds mostly in spring or early summer after soaking rains, although strong choruses sometimes form after rain at other times of the year (pers. obs.). Dry periods are spent below the surface, often under rocks or next to the foundations of walls, where soil moisture levels are higher (Minter 1997). In overcast or misty conditions, calling may continue unchecked for several days and nights (Minter 1998). Calling activity is also influenced by temperature: the chorus intensity usually drops in the early hours of the morning when temperatures are low, but increases again after sunrise, and persists until mid-morning, even on clear, hot days (pers. obs.). During the day, calling usually takes place from well-concealed, shallow depressions, but at night males often move about on the surface, calling from one location for a while before moving to another (Minter 1997, 1998).
Mozambique Rain FrogBreviceps mossambicus Near Hluhluwe, KwaZulu-Natal Photo by Nick Evans
Amplexus is by adhesion and oviposition takes place in a chamber below the soil surface. Nests excavated near Wakkerstroom were located below loose rocks on the hill slopes or under old, dry tree trunks. Females remained in the vicinity of the nests, which contained 20–25 eggs; metamorphosis was completed after 6–8 weeks (FitzSimons and Van Dam 1929; Swanepoel 1970).
After good rains, adults often emerge in large numbers from their places of concealment, to feed on alate termites (pers. obs.). At such times, many are killed on the roads.
Status and Conservation
The Mozambique Rain Frog is widely distributed in a variety of habitats and does not appear to be at risk. It occurs in a number of provincial nature reserves and national parks.
Distribution
The extralimital distribution of B. mossambicus includes southern Tanzania, Malawi, Mozambique, Zambia, Zimbabwe, and eastern Botswana (Poynton and Broadley 1985a). In the atlas region, the Mozambique Rain Frog occurs on the coastal plain of KwaZulu-Natal, extending southward as far as Mkambati (3129DA) in Eastern Cape Province. In the interior it is found in the foothills and along the crest of the Lebombo Mountains and the Great Escarpment of KwaZulu-Natal, Swaziland, Mpumalanga and Limpopo Province, extending northward as far as Haenertsburg (2329DD). The populations along the escarpment, for example, those near Wakkerstroom (2730AD), referred to B.adspersus pentheri by Poynton (1964), were assigned to B. mossambicus on the basis of advertisement call structure (Minter 1998).
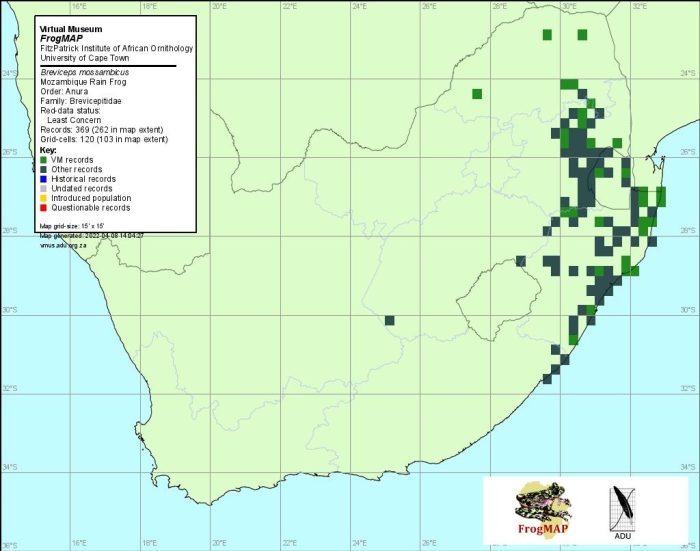
Distribution of Breviceps mossambicus. Taken from the FrogMAP database, April 2022.
The atlas data are based largely on advertisement calls and associated specimens and are considered to be reliable (Minter 1998).
Further Resources
The use of photographs by A. Coetzer, Heather Hodgson, and Nick Evans is acknowledged
Other Common Names: Mosambiekse Reënpadda (Afrikaans)
Recommended citation format: Minter, LR; Tippett, RM. (2025). Mozambique Rain FrogBreviceps mossambicus. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/16/mozambique-rain-frog-breviceps-mossambicus/
This species text has been updated and expanded from the text in the 2004 frog atlas: Minter, LR. (2004). Mozambique Rain FrogBreviceps mossambicus. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
Find the Plaintive Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
The female Plaintive Rain Frog attains a length of 53mm (males are smaller than females). The body is stout with short legs and relatively small eyes. The colouration of the upper sides is variable but is most often tan-brown to dark brown with black markings. Specimens from forested places can be mostly black, sometimes with paler spots. Both the upper and lower parts are granular.
The natural habitats of the Plaintive Rain Frog are indigenous forest and the adjacent grassland along the eastern escarpment. Along the great escarpment, the Plaintive Rain Frog occurs in Moist Upland Grassland, North-eastern Mountain Grassland, Afro Mountain Grassland, and adjacent Afromontane Forest. Along the Eastern Cape and KwaZulu-Natal coast, it inhabits Coastal Bushveld/Grassland, Coastal Forest, Sand Forest, and Dune Thicket. At Pennington in KwaZulu-Natal (3030BC), males were heard calling in dense, matted grass, five metres from the high-water mark (Minter 1998). Between the coast and the escarpment in KwaZulu-Natal, it is found in Short Mistbelt Grassland. At Eshowe, calling males were collected in suburban gardens (Minter 1998).
Coastal forest habitat. Mtunzini, KwaZulu-Natal hoto by Ryan Tippett
Factors that are common to these vegetation types, and may limit the distribution of this species, are annual rainfall in excess of 700 mm and well-shaded soil, which result in higher soil moisture levels over relatively longer periods than is the case in drier, more open habitats.
Behaviour
During dry periods, the Plaintive Rain Frog remains below the surface. Minter (1998) followed a shallow burrow for 3 m until it dipped down and disappeared under a large rock, and found deep, vertical burrows near the call sites of several males. Specimens have also been collected under rocks (Jacobsen 1989).
Plaintive Rain FrogBreviceps verrucosus Baziya forest, Eastern Cape Photo by Luke Kemp
The breeding period of the Plaintive Rain Frog is relatively short, commencing with the first spring rains in August or early September and seldom extending beyond mid-November. Choruses develop rapidly in rain, drizzle or heavy mist and continue unabated, day and night, as long as the wet weather lasts. Males are prompted to vocalize by the calls of their immediate neighbours, and in this way bouts of calling spread through the population in waves, as in the Bushveld Rain Frog B.adspersus. The Plaintive Rain Frog is easily prompted to call by a tape recording or human imitation of the call, even when no other calling is taking place (pers. obs.). Calling is inhibited by wind and low temperatures (Minter 1998).
The call site is usually a well-concealed, shallow depression under dense, matted grass or leaf litter, but males may call at the surface, or from elevated positions on grass tussocks, up to 40 cm from the ground (Poynton and Pritchard 1976; Minter 1998). Amplexus, oviposition and development have not been recorded.
Food items include amphipods, isopods, juvenile myriapods, caterpillars, beetles and ants (Poynton and Pritchard 1976).
Status and Conservation
The Plaintive Rain Frog is widely distributed in a variety of habitats and does not appear to be at risk. It occurs in a number of provincial nature reserves and in the Golden Gate Highlands National Park (Bates 1995).
Plaintive Rain Frog Breviceps verrucosus Mariepskop, Limpopo Photo by Luke Verburgt
Distribution
The Plaintive Rain Frog is endemic to South Africa, Lesotho, and eSwatini (Swaziland). It occurs along the Great Escarpment, from the Nico Malan Pass (3226DA) in Eastern Cape Province, northward through Lesotho, KwaZulu-Natal and Swaziland, to the Blyde River Canyon Nature Reserve (2430DB) in Mpumalanga. A single specimen was collected on the border of Lesotho, in eastern Free State (Bates 1995). The species descends into moist grassland and forest east of the escarpment, reaching the coast at Coffee Bay (3129CD), and extending along the coast as far north as Empangeni (2831DD). The most southerly record is based on a museum specimen from Zuurberg (3325BC).
The great variation in colour and markings present in different populations of this species is attributed to crypsis. For example, individuals living in forest and along the forest fringe at Graskop (2430DD) are black with fine, cream speckles, while those living on light sandy soils at Eshowe are uniformly light brown (Minter 1998). Populations from grassland habitats in Eastern Cape Province exhibit a varying degree of light brown mottling on a dark brown background, and were previously referred to a separate subspecies, B. v. tympanifer. However, similar marking are seen on specimens from grassland habitats in KwaZulu-Natal and Mpumalanga, while a population sampled at the grassland/forest interface at Wesa (3029DA) contains entirely melanistic as well as mottled individuals (Lambiris 1989a; Minter 1998). Therefore, B. v. tympanifer does not represent a valid subspecies, but rather a cryptic colour variation in grassland (Minter 1998). These various populations show no differences in advertisement-call structure, which confirms that they are conspecific (Minter 1998).
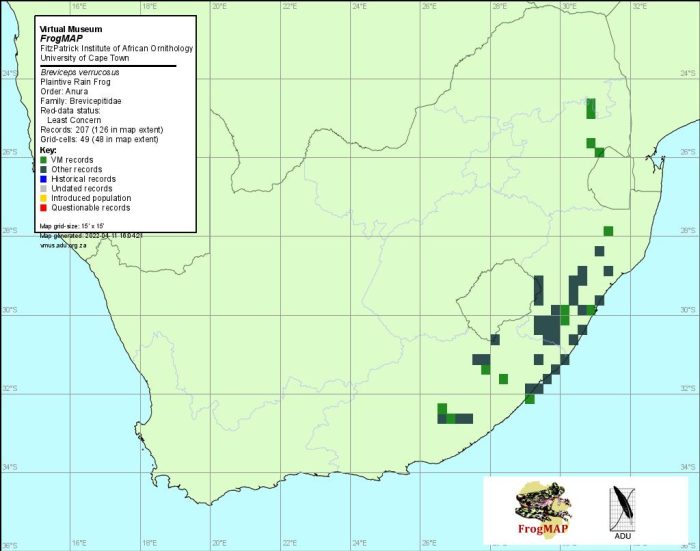
Distribution of Breviceps verrucosus. Taken from the FrogMAP database. April 2022.
Breviceps maculatus, described from montane grassland in the KwaZulu-Natal Drakensberg (FitzSimons 1947), has been recognized by some authors as a distinct species (Poynton 1964; Lambiris 1989a, 1991) on the basis of dorsal markings and larynx structure, while others have placed maculatus in the synonymy of B.verrucosus because of the similarity of their advertisement calls (Passmore and Carruthers 1979, 1995; Bates 1995; A. Channing pers. comm.; pers. obs.). B. maculatus is not treated as a separate taxon in this atlas, but an analysis of a larger sample of specimens and calls is required in order to validate this viewpoint.
The atlas data are reliable, but incomplete. The taxonomic status and distribution of populations in Lesotho, and along the escarpment in Mpumalanga, and possibly Limpopo Province, warrant further investigation (see B. sylvestris species account).
Further Resources
The use of photographs by Cornelia Rautenbach, Luke Kemp, and Luke Verburgt is acknowledged.
Recommended citation format: Minter, LR; Tippett, RM. (2025). Plaintive Rain FrogBreviceps verrucosus. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/15/plaintive-rain-frog-breviceps-verrucosus/
This species text has been updated and expanded from the text in the 2004 frog atlas: Minter, LR. (2004). Plaintive Rain FrogBreviceps verrucosus. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
View the above photo record (by Nick Evans) in FrogMAPhere.
Find the Plain Stream Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pyxicephalidae
PLAIN STREAM FROG – Strongylopus wageri
Wager, 1961
Identification
S. wageri reaches 50 mm in body length. The dorsal colouration varies from uniform light brown or yellow-green to brick red, and is slightly paler on the snout. A distinct, dark band passes from the nostril through the eye to the shoulder and then continues along the flank. Faint barring on the legs and a narrow, pale vertebral stripe may be present (Wager 1965; Lambiris 1989a).
S. wageri may be distinguished from S. grayii, which sometimes has a similar dorsal colour pattern, by the extent of the webbing on the longest toe: in S. wageri no more than three phalanges are free of webbing, whereas in S. grayii, four or occasionally three phalanges are free of webbing (Poynton 1964; Passmore and Carruthers 1995).
The advertisement call is a short cackle c.500 ms in duration, repeated at irregular intervals. A call consists of a variable number of short notes, each about 50 ms in duration (Passmore and Carruthers 1995; Channing 2001).
Wager (1986) noted that tadpoles from Weza, Giant’s Castle and Mont-aux-Sources were identical, but Lambiris (1989a) found that adults from both Giant’s Castle Game Reserve and Entumeni Nature Reserve differ from the type series (Weza) with regard to webbing and colour pattern and suggested that they may represent an undescribed sibling species. The taxonomic status of the various isolated populations that do not form part of the main KwaZulu-Natal Drakensberg population, should also be examined as they may be in the process of speciation.
Habitat
At lower altitudes, S. wageri inhabits mistbelt forest, while at high altitudes up to 2000 m, it occurs in montane grassland. Adults may be found amongst vegetation or stones on the banks of clear streams and pools. Breeding takes place in clear, cold, fast-flowing streams and their pools.
Behaviour
Adults are active in the late afternoon and at night. When disturbed they jump into the water and remain motionless in the mud at the bottom, apparently relying on camouflage to avoid detection.
S. wageri breeds January–May, but mainly in autumn when water levels have stabilized (Channing 1979). Males call from concealed or partly concealed positions along the edges of streams or pools and sometimes under water (Lambiris 1989a).
In the laboratory, spawning was recorded at about 03:00. Clutches of 12–250 eggs are firmly attached to twigs or blades of grass dangling or submerged in the water, or to submerged rocks. The eggs measure 2.8 mm in diameter and are enclosed in a stiff, jelly-like capsule with a diameter of 5 mm, that swells to 7 mm after a few days. Eggs hatch after 8–9 days (Wager 1986; Channing 1979).
The tadpoles lie in the algae and mud at the bottom of the pools, moving slowly, but darting around rapidly if disturbed. They are often found in sunny positions in shallow water (Channing 1998). Tadpoles grow slowly in the cold of winter, and metamorphosis is completed after 5–9 months.
Adult S. wageri prey on terrestrial arthropods and gastropods (Channing 1979), as well as a variety of aquatic and semi-aquatic insects (Lambiris 1988e). In turn, they are probably eaten by a variety of vertebrates such as birds, snakes, otters and other frogs. Channing (1979) observed a crab climbing onto vegetation to devour the eggs of this species.
Status and Conservation
Status
S. wageri is placed in the category Near Threatened (Harrison et al. 2001; this publication) because of its small extent of occurrence and area of occupancy, fragmented distribution, and an existing and projected decline in the extent and quality of its habitat over a large part of its range. The species was not placed in a threatened category because it is relatively well protected in the reserves along the KwaZulu-Natal Drakensberg. It should be noted that S. wageri is heavily dependent on the continued protection of its habitat in these areas.
S. wageri is recorded from several conservation areas in KwaZulu-Natal, namely Ukahlamba Drakensberg Mountain Park, as well as Entumeni Nature Reserve, Ntsikeni Nature Reserve, Qudeni State Forest, Ngele Forest Nature Reserve and Ngome Forest Reserve.
Threats
In the past, S. wageri was probably more widespread on the Drakensberg escarpment in Mpumalanga, than at present. Much upland habitat in these areas has been destroyed by intensive afforestation with alien trees, and this is likely to have affected local populations of S.wageri. Channing (1979) noted that the destruction of indigenous forests has probably also resulted in loss of habitat to this species in KwaZulu-Natal.
Habitat loss to S. wageri is estimated to be >20% over the past 50 years, with a similar rate of decline in population numbers due to afforestation, chemical pollution associated with sylviculture, alien-plant invasions and the introduction of trout into streams (Harrison et al. 2001).
Recommended conservation actions
Extensive distribution surveys and monitoring of known populations are recommended. Habitat management, including the identification of limiting factors, is needed for effective conservation of this species (Harrison et al. 2001). It is especially important to determine whether S. wageri is able to survive in streams that run through plantations.
Distribution
S. wageri is endemic to South Africa. It has not been recorded at or near the summit of the Drakensberg in Lesotho (Bates and Haacke 2003). Distribution records are sparse and widely scattered. From the type locality on the Ngele Mountains near Weza Forest (3029DA) in KwaZulu-Natal, the species ranges northward along the escarpment and foothills of the Drakensberg mountain range. Isolated populations exist further to the east in KwaZulu-Natal, at Qudeni Forest (2830DB), Entumeni Nature Reserve (2831CD) and Ngome Forest Reserve (2731CD), and along the southern boundary of Mpumalanga Province.
Acocks (1975) suggested that in the recent past (A.D. 1400) KwaZulu-Natal was covered by forest and scrubforest, with grassveld on the summit of the Drakensberg. If so, this may explain the present disjunct distribution of this species in patches of forest that were once connected (Channing 1979).
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Wager se langtoonpadda (Afrikaans)
Recommended citation format for this species text:
Bates MF, Tippett RM. Plain Stream Frog Strongylopus wageri. BDI, Cape Town. Available online at http://thebdi.org/2021/12/15/plain-stream-frog-strongylopus-wageri/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Bates MF 2004 Strongylopus wageri Plain Stream Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Felicity Grundlingh) in FrogMAPhere.
Find the Cape Platanna in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pipidae
CAPE PLATANNA – Xenopus gilli
Rose & Hewitt, 1927
Identification
X. gilli is a typical member of the genus Xenopus. The upper body is light to yellow-brown with elongated, dark brown patches, sometimes paired, that begin between the eyes and extend backwards, breaking up into smaller patches on the lower back and upper surfaces of the hind limbs. The underside usually has clear blackish and yellow mottling, but this may be pale and indistinct in some individuals, especially in populations near Kleinmond (3419AC; J.A. Harrison in litt.).
X. gilli can be distinguished from X. laevis by the following morphological features: its smaller size (<60 mm in body length); a narrower, more acutely pointed head; the absence of a subocular tentacle (present but inconspicuous in laevis); a poorly developed inner metatarsal tubercle (a distinct ridge in laevis); and less conspicuous lateral line sense organs (Poynton 1964; Picker and de Villiers 1988; Passmore and Carruthers, 1995; Kobel et al. 1996; Channing 2001).
The advertisement call consists of a series of short, rapidly pulsed, metallic buzzes emitted under water at a rate of about two per second (for further details of call structure see Passmore and Carruthers 1995; Picker et al. 1996; Channing 2001).
Habitat
X. gilli inhabits blackwater wetlands in low-lying coastal areas. These are permanent and seasonal seepages, marshes, ponds, pans, vleis and coastal lakelets, in a variety of fynbos vegetation types and, in places, a mixture of fynbos and dune thicket. The vegetation types include mostly Mountain Fynbos, Sand Plain Fynbos (on the Cape Flats), or Mountain Fynbos mixed with either Limestone Fynbos or Dune Thicket. The substrate has a predominantly sandy base and varies, depending on the humic content, from white or grey to a dark brown or blackish soil.
The water is humic and dark in colour, low in nutrients, high in dissolved solids, and typically has a low pH (minimum 3.4; Picker 1985). It has been demonstrated that the tadpoles of X. gilli can tolerate pH as low as 3.6, whereas X. laevis tadpoles have a reduced rate of survival below pH 5–6. This accounts for the observed habitat segregation between these species on the Cape Peninsula where X. gilli occurs in acidic blackwater seepages and ponds, while X. laevis prefers clear water bodies with elevated pH, for example, artificial impoundments (Picker et al. 1996). Disturbances of X. gilli water bodies which alter the humic content and nutrient levels, cause an increase in pH levels and often result in the colonisation of these water bodies by X. laevis, providing an opportunity for hybridization between the species (Simmonds 1985; Picker et al. 1996).
While X. gilli avoids habitats that have been disturbed by urban development or agriculture, or that contain invasive plants and animals (Picker 1985; Picker and De Villiers 1989), it is interesting to note that healthy populations of this frog inhabit certain seepages that were excavated to form waterholes in the Cape of Good Hope area of the Cape Peninsula National Park (CPNP). However, no other habitat disturbances or threats are evident in these pools, the water quality meets the necessary requirements, and consequently X. gilli populations have flourished in them for well over two decades.
Behaviour
When its wetland habitat dries up during the summer months, X. gilli survives by aestivating below the surface. Rau (1978) found several specimens encapsulated in the mud of dried-up vleis on the Cape Flats, and Rose (1962: 33) found one individual at a depth of 15 cm amongst the roots of a large “weed” growing on the site of a small dried-up vlei. During the rainy season, overland migrations between ponds have been observed in Cape Peninsula National Park (Picker 1985).
Breeding commences during the wet winter months (July), and continues until late October (Rau 1978). Three to four hundred dark brown eggs are laid over a period of a day, each surrounded by a jelly capsule 1.3 mm in diameter (Channing 2001). The nektonic tadpoles feed on phytoplankton in the water and complete their metamorphosis by the end of summer. Rau (1978) recorded spawning activity over a four-month period and found metamorphosis to take about 120 days. The breeding season of X. gilli overlaps that of X. laevis, which increases the opportunity for hybridization (Rau 1978; Picker et al. 1996).
Adult frogs feed on living and dead animal material in their wetland habitat, including aquatic invertebrates and the eggs, tadpoles and smaller frogs of their own kind and other species (Picker and De Villiers 1988). Predation of the immature stages of X. gilli by the larger X. laevis is presumably intense in disturbed habitats that have been invaded by the latter. Other predators include herons, cormorants and water mongoose.
Status and Conservation
Status
Development and general habitat degradation have severely impacted on the extent of occurrence and area of occupancy of this species, resulting in a loss of more than 50% of its habitat, and severe fragmentation of its populations (Harrison et al. 2001). This is particularly serious on the Cape Flats and adjoining Cape Peninsula where extensive urban development has taken place. By now, X. gilli is possibly extinct in both of these areas except for populations in the Cape Peninsula National Park. Similar threats are escalating between Rooiels and Kleinmond on the south coast, leading to further habitat loss and fragmentation.
About 70% of all currently known X. gilli habitat is situated in the Cape Peninsula National Park. This is the stronghold of the species with healthy populations in the Cape of Good Hope area. The populations at some sites in this area have been found to vary from 121–591 frogs (Picker and De Villiers 1989; Picker et al. 1996). It is nevertheless of concern that X. laevis and/or hybrids have been reported from most of the wetlands in this area, and the situation needs to be monitored. It is also of concern that the only X. gilli habitat protected within a conservation area, besides Cape Peninsula National Park and Greater Betty’s Bay Nature Reserve, is a small remnant of habitat in Agulhas National Park.
X. gilli was included in the first South African Red Data book for amphibians, in the Rare category (McLachlan 1978). In the revision (Branch 1988), it was classified Endangered. Endangered status was retained in Harrison et al. (2001), based on an extent of occurrence <5000 km2, an area of occupancy <500 km2, a severely fragmented habitat, continuing decline in the extent of occurrence, area of occupancy, extent and quality of habitat and the number of locations/subpopulations and mature individuals. The species is legally protected by Nature Conservation Ordinance 19 of 1974, but is not listed by CITES.
Threats
Loss of habitat and habitat fragmentation pose the most serious threats to the survival of X. gilli. In particular, urban development and agriculture have resulted in the filling in and drainage of its wetland habitat or have led to the pollution and eutrophication of breeding sites. Furthermore, the building of artificial reservoirs and irrigation systems has enabled the highly adaptable X.laevis to invade areas from which it was previously excluded, including disturbed blackwater wetlands containing populations of X. gilli. Predation by X. laevis on the eggs, tadpoles and froglets of X. gilli represents a further threat to the survival of X. gilli.
Urban expansion and human activities also accelerate the spread of invasive alien vegetation. At some localities, indigenous fynbos vegetation has been replaced by stands of exotic trees (Port Jackson Willow Acacia saligna and Rooikrans A. cyclops). This alters the water chemistry and results in unsuitable habitat for X. gilli. Introduced predatory fish may pose an additional threat, particularly in some of the larger, permanent wetlands.
The invasion of disturbed X. gilli habitat by X. laevis, and the subsequent hybridization of these two species, has been well documented (Rau 1978; Picker 1985; Simmonds 1985; Picker and De Villiers 1989; Picker et al. 1996; Evans et al. 1998). Hybridization threatens the gene pool of the smaller, less numerous X. gilli, through potential genetic swamping of populations. This threat is present throughout the distribution area of X. gilli (Picker et al. 1996) but appears not to be as serious as was previously thought (Evans et al. 1998).
Recommended conservation actions
The distribution and conservation status of X. gilli is monitored by the Western Cape Nature Conservation Board (WCNCB) as part of a threatened species monitoring programme (De Villiers 1997). Healthy populations in the Cape of Good Hope area were monitored mainly by the Zoology Department of the University of Cape Town, but monitoring ceased in 2000. It will be continued by the CPNP in conjunction with the WCNCB. Besides the monitoring of known localities, additional survey work is to be conducted in surrounding areas, including montane habitats.
Although X. gilli populations on the Cape Peninsula are well protected, it is important that other viable populations be included in statutory conservation areas and managed appropriately. This was strongly recommended by Evans et al. (1997), who found that populations to the east of False Bay showed significant genetic differences from the Cape Peninsula populations, although these were not considered to be taxonomically significant. In particular, they indicated that “protective measures within X. gilli habitat near Kleinmond would conserve much of the genetic diversity seen in this species”.
The main management activity is control of alien vegetation. This is undertaken on the Cape Peninsula by CPNP, in the Kleinmond area by the WCNCB, and in the Betty’s Bay area by the Overstrand Municipality. Alien vegetation clearing programmes are now underway in the Gansbaai to Agulhas area, but they need to be intensified in X. gilli habitat.
In 1985, a Cape Platanna Conservation Committee built a precast wall around Geps Dam, one of the pools in the Cape of Good Hope area, to protect its X. gilli population from contamination with X. laevis (Picker and de Villiers 1989). After the wall was built, X. laevis and hybrids were removed from the water body. Holes were regularly found under the wall, but no X. laevis or hybrids were observed in the water body when it was last examined in 2000. In fact, X. laevis and hybrids are apparently on the decline in at least this section of the Cape of Good Hope area (M.D. Picker pers. comm.). Although the situation requires continued monitoring, it seems that the wall now serves little purpose and might safely be removed.
In 1988, the Cape Platanna Conservation Committee translocated 154 juvenile X. gilli from the Cape of Good Hope area to four blackwater pools in the Silvermine nature area, both areas falling within the present CPNP. Although a survey produced no sign of X. gilli in the Silvermine nature area shortly before the translocation, about 16 specimens were collected in 1926 from somewhere in and next to the “Sylvermyn River” which drains this area (Rose and Hewitt 1927). The main reason for the translocation was to establish a separate breeding colony of X. gilli on the Cape Peninsula that would perhaps be free of the X. laevis threat. It would appear that this experiment has had some success: from one to six adults have been seen in one of the pools on about four occasions during the 10-year period following the translocation. Further recommendations are being formulated in this regard, and monitoring work is to continue.
Distribution
This species is endemic to the winter rainfall region of the Western Cape, generally occurring in relatively low-lying areas (10–140 m a.s.l.) within 10 km of the coastline. Its distribution is correlated with the presence of nearby mountain ranges and an annual rainfall exceeding 500 mm p.a. Confirmed records span a distance of about 160 km, from the Cape Peninsula southeastward towards the Agulhas district (Picker and de Villiers 1988, 1989).
For about 40 years following its description in 1927, the species was known only from the Cape Peninsula and adjoining Cape Flats (3418AB, BA; 3318CD, DC). However, in the late 1960s it was discovered further south on the Cape Peninsula in the Cape of Good Hope Nature Reserve (3418AD), and in the period from 1973 to 1988 it was collected along the coastal forelands to the southeast of the Cape Peninsula and Cape Flats, at Hangklip and Betty’s Bay (3418BD), Kleinmond (3419AC), midway between Gansbaai and Agulhas (3419DA), and northwest of Agulhas at the eastern base of Soetanysberg (3419DB, DD).
Two inland records from Nieuwoudtville (3119AC) in 1898 and Citrusdal (3219CA) dating from about 1937 (Rau 1978), have not been subsequently confirmed despite several field trips to these areas. The validity of these records is in doubt and they are excluded from the map.
During the 1980s a study of the distribution and habitat requirements of X. gilli was undertaken to assess its future survival prospects. The study showed that, although the species still occurred in at least 24 localities in seven quarter-degree grid cells, habitat destruction and degradation had led to extinction at 60% of its known localities (Picker and De Villiers 1989).
Thus,X. gilli has been recorded from a total of 10 quarter-degree grid cells (excluding the two inland localities), but since 1995 it has been found in only five of these cells (3418AB, AD, BD; 3419AC, DA). The atlas data are reliable.
Distribution of Xenopus gilli. Taken from the FrogMAP database as at November 2021.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gill’s Platanna (Alternative Common Name); Kaapse Platanna (Afrikaans)
Recommended citation format for this species text:
de Villiers AL, Tippett RM. Cape Platanna Xenopus gilli. BDI, Cape Town. Available online at http://thebdi.org/2021/12/15/cape-platanna-xenopus-gilli/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
de Villiers AL 2004 Xenopus gilli Cape Platanna. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.







































{kind=link}
{kind=link}
{kind=link}
{kind=link}