View the above photo record (by Faansie Peacock) in FrogMAPhere.
Find the Plain Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
The Plain Rain Frog is a stout species with short legs and small, dark eyes. Females can attain 51mm in length and are larger than the males. The upper sides are dark brown to black and devoid of markings. The underside is smooth and dark, sometimes with indistinct light spots.
Habitat
The Plain Rain Frog inhabits Afromontane Forest and Mountain Fynbos along the southern coastal belt (Burger 1992; D. McDonald pers. comm.). It occurs at a range of altitudes to well in excess of 1000 m above sea level. The Plain Rain Frog also occurs down to sea level, where mountains reach the coast. The Grootvadersbosch area is transitional between winter and all-year rainfall, with an average annual rainfall of 1050 mm. Relatively dry periods are May–July and December–January. The forests at Grootvadersbosch are situated on clay soils derived from Bokkeveld shales, while the fynbos occurs on sandy soils derived from Table Mountain Sandstone.
Behaviour
Plain Rain FrogBreviceps fuscus Garden Route Botanical Garden, Western Cape Photo by Christiaan Steenkamp
The Plain Rain Frog often occurs at high densities in suitable habitats. In forest these frogs may be found in short, vertical tunnels under logs (pers. obs.). When disturbed, they inflate their bodies to such an extent that they are difficult to remove from the tunnels – probably an adaptive strategy to escape predation. The vertical tunnels are connected to a network of shallow horizontal tunnels leading away in all directions, from which they emerge to feed (L.R. Minter pers. obs.).
The Plain Rain Frog is a summer breeder that starts to call in September (H. Braack pers. comm.), forming large choruses that may continue for several days in rainy weather. Males may call from vegetation up to 1 m above the surface, (pers. obs.; D. McDonald pers. comm.) and sometimes from within their shallow burrows.
Adhesive amplexus occurs (Visser et al. 1982). The eggs are 5 mm in diameter, within 8 mm capsules, and yellow in colour. The egg chambers are c.30–40 mm below the surface and the known clutch sizes are 42 and 43 eggs, each topped with a layer of 25–30 empty egg cases (Channing 2001).
Predation by the Bush Pig Potamochoerus larvatus has been recorded, with 18 B. fuscus specimens found in the stomach of one pig (Palmer 1982).
Status and Conservation
Although the Plain Rain Frog has a relatively restricted distribution range, it appears not to be of conservation concern. Overutilisation of indigenous forests and the creation of exotic timber plantations during the past century or more undoubtedly impacted some populations, but the species is still abundant at several localities. It is known from the following protected areas: Tsitsikamma National Park (Carruthers and Robinson 1977; Branch and Hanekom 1987), Outeniqua Conservation Area, Grootvadersbosch Nature Reserve, and Marloth Nature Reserve.
Plain Rain FrogBreviceps fuscus Saasveld, Western Cape Photo by Damian van Aswegen
Distribution
The Plain Rain Frog is endemic to the southern Cape Fold Mountains in the Western and Eastern Cape provinces of South Africa. It occupies a narrow, linear strip along the south-facing slopes of the mountain ranges that run parallel to the coast, from the Langeberg Mountains near Swellendam (3320CD) in the west, to Dieprivier Forest near Kareedouw Pass (3324DC) in the east.
Distribution of Breviceps fuscus. Taken from the FrogMAP database, April 2022.
An aural record from Zuurberg (3325BC; H.H. Braack pers. comm.) is not shown on the map and requires confirmation. B.fuscus is sympatric with B.acutirostris at Grootvadersbosch Nature Reserve (3320DD) and adjacent grid cells, and with B. montanus over most of its range. The distribution pattern of this species is, for the most part, continuous except for a break between the Langeberg and Outeniqua mountains. Speculation that B. fuscus might occur at Joubertina, Niekerksberg, and Prince Alfred Pass (Burger 1992) has not been confirmed. Calls heard at Joubertina were subsequently attributed to B.adspersus pentheri (pers. obs.), and the other records require verification.
Further Resources
The use of photographs by Christiaan Steenkamp, Damian van Aswegen, and Faansie Peacock is acknowledged.
Plain Rain FrogBreviceps fuscusHewitt, 1925
Other Common Names: Black Rain Frog Frog (Alt. English); Gewone Janblom (Afrikaans)
Recommended citation format: Burger, M; Tippett, RM. (2025). Plain Rain Frog Breviceps fuscus. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2022/01/25/plain-rain-frog-breviceps-fuscus/
This species text has been updated and expanded from the text in the 2004 frog atlas: Burger, M. (2004). Plain Rain Frog Breviceps fuscus. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
Find the Bushveld Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
Bushveld Rain FrogBreviceps adspersus Near Hoedspruit, Limpopo Photo by Alison Sharp
The Bushveld Rain Frog is a medium-sized species. Males attain 47mm in length, while females can reach 60mm. The limbs are stubby and the head is short. The dorsal surface is rough and granular, with variable colourations and patterning, but is most often brown with paired paler, yellowish patches. Variable darker markings are also present, including a blackish stripe running from the eye toward the armpit. The undersides are smooth and white with some darker mottling and two black patches on the throat.
Habitat
The Bushveld Rain Frog inhabits seasonally dry to semi-arid habitats in the Savanna Biome, with sandy to sandy-loam soils. It is particularly associated with bushveld vegetation types that are characterised by “a grassy ground layer and a distinct upper layer of woody plants” (Low and Rebelo 1996). It is conspicuously absent from the Grassland and Forest biomes.
The subspecies B. a. pentheri occurs in Eastern Thorn Bushveld, Spekboom Succulent Thicket, and Valley Thicket. In Polokwane and Grahamstown, parks and gardens with well-turned, sandy soil contain breeding populations of both B. a. adspersus and B. a. pentheri, indicating that this species can survive in a suburban environment.
Habitat at Mokala National Park, Northern Cape Photo by Karis Daniel
Behaviour
Bushveld Rain Frogs spend the winter dry season 15–30 cm below the surface, often in situations where soil moisture is conserved, for example, next to or under rocks, logs, stumps, or tree roots (Jacobsen 1989; pers. obs.).
In spring or early summer, following heavy rain, male Bushveld Rain Frogs emerge from the soil and establish call sites 5–200 cm from their winter retreats. The call site usually consists of a well-concealed shallow depression, about the depth of the frog’s body, at the base of a grass tuft or small herbaceous plant. In overcast, damp conditions, calling may continue unchecked for several days and nights. Males are prompted to call by the calls of their immediate neighbours, and this results in bouts of calling which spread through the population in waves. When hot, dry weather returns, or when disturbed, males retreat to their underground burrows. Males were observed to use the same call site for up to five consecutive nights (Minter 1995, 1998).
Amplexus is facilitated by a sticky skin secretion which ensures that the male remains attached to the female during nest construction. A mass of about 45 eggs, covered by a smaller mass of fluid-filled jelly capsules lacking yolk, is deposited in a chamber about 30 cm below the surface. The female remains nearby until the froglets are ready to leave the nest (approximately six weeks). The reason for her presence has not been established (Minter 1995, 1998).
The Bushveld Rain Frog does not appear to be at risk as much of its habitat is used for game and cattle farming, and is relatively undisturbed. It also occurs in a number of provincial nature reserves and national parks.
Distribution
Beyond the atlas region, the Bushveld Rain Frog occurs in Namibia, Botswana, Zimbabwe, southern Zambia, and Mozambique (Poynton and Broadley 1985a). In the atlas region, the subspecies B. a. adspersus occurs in its preferred habitat throughout most of Limpopo and North West provinces, the northern Free State, the eastern parts of the Northern Cape, Gauteng, Mpumalanga, and central and eastern eSwatini (Swaziland). In KwaZulu-Natal, it is found at lower elevations, as far south as Margate (3030CD).
Call data and molecular evidence (Minter 1998; Engelbrecht and Mulder 2000) support the retention of the subspecies B. a. pentheri in the Eastern Cape, where it is distributed from Joubertina (3323DD) inland to Grahamstown (3326BC), and northward as far as the Stutterheim district (3227BC). Populations from the Great Escarpment of KwaZulu-Natal and Mpumalanga, e.g., near Wakkerstroom (2730AD), previously referred to B.a.pentheri by Poynton (1964), were assigned to B.mossambicus on the basis of advertisement call structure (Minter 1998).
Distribution of Breviceps adspersus. Taken from the FrogMAP database, April 2022.
Light paravertebral and dorsolateral patches on the dorsum of the Bushveld Rain Frog distinguish this species from northern Mozambique populations of B. mossambicus, which lack these markings (Poynton 1964; Poynton and Broadley 1985a; Lambiris 1989a; Minter 1998). However, this is not the case in the atlas region, where similar markings are present in B. sopranus, B. bagginsi, and in populations of B. mossambicus from the Great Escarpment and coastal KwaZulu-Natal (see B. mossambicus species account).
A study of advertisement calls (Minter 1998) did not support the hypothesis that B. adspersus and B.mossambicus hybridize extensively in KwaZulu-Natal (Poynton 1964; Poynton and Broadley 1985a; Lambiris 1989a). The hybridization hypothesis was based on dorsal markings that do not constitute a reliable diagnostic character. Therefore, existing museum and literature records of B. adspersus from KwaZulu-Natal, unless based on advertisement calls, should be regarded as unreliable. A survey based on advertisement calls is needed in order to re-map the distribution of B. adspersus in the northeastern parts of the atlas region, and the atlas data from the pre-1996 period should be viewed with circumspection.
Further Resources
The use of photographs by Alison Sharp, Kyle Finn, Karis Daniel, and Vaughan Jessnitz is acknowledged.
Bushveld Rain FrogBreviceps adspersusPeters, 1882
Other Common Names: Common Rain Frog (Alt. English); Bosveld Reenpadda (Afrikaans)
Recommended citation format: Minter, LR; Tippett, RM. (2025). Bushveld Rain Frog Breviceps adspersus. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2022/01/24/bushveld-rain-frog-breviceps-adspersus/
This species text has been updated and expanded from the text in the 2004 frog atlas: Minter, LR. (2004). Bushveld Rain Frog Breviceps adspersus. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
Find the Strawberry Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
Strawberry Rain FrogBreviceps acutirostris Landdroskop, Western Cape Photo by Felicity Grundlingh
The female Strawberry Rain Frog attains a maximum length of 40mm. Males are much smaller, reaching just 25mm in length. The body is stout with short legs, and the snout is slightly more pointed than in other Breviceps species. The dorsal surface is reddish or pink to creamy white with many elevated black granules. The undersides are purplish with cream-coloured spots.
Habitat
The Strawberry Rainfrog occurs only in areas of high rainfall in uplands and mountains, both in Mountain Fynbos and in Afromontane Forest. Where mountains reach the coast, the species can occur at sea level (e.g. Betty’s Bay 3418BD; Grootbos 3419CB). The soils in these habitats are coarse, acidic sands derived from Table Mountain Sandstone, frequently admixed with peaty deposits.
Behaviour
Calling occurs both at night and during the day, usually during and after rain showers in winter and spring (June–November), but dense mist may be sufficient to stimulate calling. In Grootvadersbosch forest, males have been observed calling from above the ground, on top of logs (C.N. Spottiswoode pers. comm.; H. Braack pers. comm). In fynbos, the species has been found calling above ground in dense vegetation (M. Burger pers. comm), and from short burrows situated below low, covering vegetation (pers. obs.). Adhesive amplexus is employed during mating.
An amplexed pair, with a freshly laid egg mass comprising 24 eggs, was unearthed in Mountain Fynbos in the Boosmansbos Wilderness Area, on 22 October 1986 (D. McDonald pers. comm.). The pair was found in a chamber at the lower end of a network of tunnels covering an area of about 1 m2. Some branches of the tunnels ended blindly while others formed loops off the main tunnel leading to the egg chamber. The nest site was located on a cool, moist slope in sandy loam soil, covered by prostrate Cliffortia ferruginea. The eggs measured 6.5–7.8 mm (mean = 7) in diameter (L.R. Minter unpubl. data).
Strawberry Rain FrogBreviceps acutirostris Grootvadersbosch, Western Cape Photo by Trevor Hardaker
Little is known about the biology of this species. Studies of the differences in call structure and habitat partitioning between B. acutirostris and B. fuscus, in areas of sympatry, would be of particular interest.
Status and Conservation
The montane habitat of the Strawberry Rain Frog is generally little disturbed and occurs within a number of protected areas such as the Marloth Nature Reserve, Boosmansbos Wilderness Area, and Grootvadersbosch State Forest. This species is not threatened.
Distribution
The Strawberry Rain Frog is restricted to the southwestern ranges of the Cape Fold Mountains. The most westerly range in which it occurs is the Hottentotsholland Mountains (3318DD, 3418BB), the most easterly the Langeberg Mountains (3321CD); these ranges also represent the northern limits of the species. The map indicates a gap in distribution corresponding to the eastern Hottentotsholland and Riviersonderend mountains; this apparent absence requires confirmation. The species does not occur on the Cape Peninsula. The eastern extremities of its range overlap that of B. fuscus.
Distribution of Breviceps acutirostris. Taken from the FrogMAP database, April 2022.
The atlas records are reliable, but sparse. Additional surveys may fill some of the gaps, especially in mountainous areas not adequately covered during the atlas project. The advertisement calls of B. acutirostris and B.fuscus are quite similar in structure (Passmore and Carruthers 1995; Channing 2001) although the call rate is distinctly faster in acutirostris (L.R. Minter pers. obs). In areas where both species may occur, aural records should have been supported by tape recordings of the calls, and by voucher specimens. This was seldom done, resulting in the possibility of some misidentifications.
Further Resources
The use of photographs by J. Heymans, Felicity Grundlingh, and Trevor Hardaker is acknowledged.
Other Common Names: Cape Short-headed Frog (Alt. English); Aarbei-blaasoppadda (Afrikaans)
Recommended citation format: Harrison, JA; Tippett, RM. (2025). Strawberry Rain Frog Breviceps acutirostris. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2022/01/24/strawberry-rain-frog-breviceps-acutirostris/
This species text has been updated and expanded from the text in the 2004 frog atlas: Harrison, JA. (2004). Strawberry Rain Frog Breviceps acutirostris. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
View the above photo record (by Cornelia Rautenbach) in FrogMAPhere.
Find the Guinea Shovel-nosed Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Hemisotidae
GUINEA SHOVEL-NOSED FROG –Hemisus guineensis
Cope, 1865
Habitat
Channing (2001: 122) recorded the habitat of H. guineensis as “grassland and open bush where temporary pans are formed in the rainy season”. The recorded localities for H. g. broadleyi in Limpopo Province are situated in Mopane Bushveld in the Savanna Biome.
Behaviour
Little is known of the biology of this frog. Its breeding behaviour is probably similar to that of the other species in the genus.
In the atlas region, H. guineensis has a peripheral distribution. Although it is not known to occur in any protected areas, the areas from which it has been recorded do not appear to be seriously threatened by habitat loss or degradation.
Distribution
H. guineensis broadleyi, a subspecies of the West African H. guineensis, is distributed through Angola, northern Botswana, Zimbabwe, Zambia and Mozambique.
Museum specimens collected at two localities in Limpopo Province: the farms Bridgewater (263MS; 2229CA), 25 km northeast of Alldays, and Rochdale (700MS; 2229DC), 8 km east of Waterpoort, have been assigned to this species (Poynton and Broadley 1985a; Jacobsen 1989). However, Poynton and Broadley (1985a) experienced difficulty in separating some specimens of H.g. broadleyi from H. marmoratus, using the morphological criteria identified by Laurent (1972). They suggested that in this area a complex situation exists, involving intergradation between the two taxa, and noted that the problem could only be resolved by the collection of field data and a larger series of specimens.
Similarly, Channing and Griffin (1993) reported an extreme range of morphological variation in a sample of 50 Hemisus specimens collected in the Caprivi, on the basis of which some individuals could be assigned to H. marmoratus and others to H. guineensis microps. However, only one call type was heard during three months of field work, and they concluded that only H. marmoratus was present in the Caprivi.
Channing’s (2001) description of the advertisement calls of H. marmoratus and H. g. broadleyi indicates a difference of 1000 Hz in emphasized frequency, while the pulse rate of H. g. broadleyi is approximately half that of H. marmoratus. When this part of Limpopo Province was surveyed during the atlas period, no calls were heard that could be attributed to H. g. broadleyi or to intergrades. H. marmoratus was heard calling at a number of localities in Limpopo Province, but the calls did not differ from those of H. marmoratus from other parts of the atlas region (pers. obs).
Thus it seems that H. g. broadleyi, if present at all, must occur in very low numbers in the atlas region. This is an open question that invites further investigation in the form of more intensive surveys in the area between the Soutpansberg Mountains and the Limpopo River.
No map is currently available for this species.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Guinea Snout-burrower (Alternative English Name); Guinease Graafneuspadda (Afrikaans)
Recommended citation format for this species text:
Minter LR, Tippett RM. Guinea Shovel-nosed Frog Hemisus guineensis. BDI, Cape Town. Available online at http://thebdi.org/2022/01/23/guinea-shovel-nosed-frog-hemisus-guineensis/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Minter LR 2004 Hemisus guineensis Guinea Shovel-nosed Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Jean Paul Brouard) in FrogMAPhere.
Find the Natal Leaf-folding Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Hyperoliidae
NATAL LEAF-FOLDING FROG – Afrixalus spinifrons
(Cope, 1862)
Identification
This species belongs to the dwarf Afrixalus complex, whose members are difficult to distinguish from one another morphologically, but whose advertisement calls are diagnostic (Backwell and Passmore 1991). A.spinifrons may be distiguished from sympatric Hyperolius species by the presence of a vertical pupil and tiny asperities on the skin.
The distribution and structure of the asperities is variable. In 60% of A. s. spinifrons males, there is a concentration of tubercular asperities on the snout, that is moderately to markedly swollen, almost bulbous. Along the Eastern Cape and KwaZulu-Natal coast, this character may be used to distinguish A. spinifrons from the other sympatric dwarf Afrixalus species, namely A.delicatus and A. aureus. However, at higher altitudes, populations assigned to A. s. intermedius lack a swollen snout and the asperities are distributed evenly over the dorsum (Pickersgill 1984, 1996, 2000).
In A. s. spinifrons, the dorsum is ivory to golden yellow with a broad, brown, median wedge that starts at the tip of the snout and broadens posteriorly, where it is split in two by a narrow light-coloured wedge over the urostyle. In A. s. intermedius, these dark markings are absent or confined to the posterior part of the dorsum. The light to dark, lateral band present in all dwarf Afrixalus species, is unmarked in A. spinifrons but speckled to a varying extent in A. aureus and A. delicatus. A dark, transverse band on the tibia distinguishes A. aureus from A. spinifrons. (Pickersgill 1984, 1996, 2000; Passmore and Carruthers 1995).
The advertisement calls of A. spinifrons and A. delicatus consist of two components, a short “zip” and a prolonged “trill”, while A. aureus produces only the trill component. It has been demonstrated that the trill component functions in female attraction in A. delicatus and A. aureus, and that females only respond to calls of their own species. A similar function is presumed for A.spinifrons (Backwell and Passmore 1991).
In A. s. spinifrons, the trill component has a lower emphasized frequency and a higher pulse rate than that of A.delicatus (Backwell and Passmore 1991; Passmore and Carruthers 1995; Pickersgill 1996). The published call data are based on small samples and exhibit a wide range of variation. Further investigation of the differences in advertisement call within the A. spinifrons complex is needed (see also comments under A. knysnae).
A. spinifrons bears a close resemblance to A. knysnae, but is slightly smaller (16.4–24.6 mm versus 22.0–25.2 mm) and has a larger gular disc (Pickersgill 1996, 2000). The ranges of the two species are separated by about 400 km.
A. spinifrons inhabits Coastal Bushveld-Grassland and Moist Upland Grassland. Armstrong (2001), using the Biosource Programme, modelled the potential distribution range of A. s. intermedius to include these vegetation types: Moist Coast Hinterland Ngongoni Veld, Moist Midlands Mistbelt, Moist Highland Sour Veld and Dry Highland Sour Veld (Harrison et al. 2001). In low-lying areas along the coast, A. s. spinifrons breeds in standing water, in dense sedge beds and inundated, grassy wetlands with abundant surface vegetation, while at higher altitudes, it inhabits marshes, dams, floodplains and riverbanks (Lambiris 1989a; Pickersgill 1996).
Behaviour
During the day, both subspecies retreat into the leaf axils of grasses, rushes and arum lilies Zantedeschia spp., particularly those standing in or immediately adjacent to water, and A. s. intermedius may also be found sunbathing in arum lily blooms. This habit is adopted almost exclusively by females and juveniles (Pickersgill 1996). In winter, A. s. spinifrons is often encountered in the leaf axils of Strelitzia and bananas.
Breeding activity has been reported August–February for A. s. spinifrons, and September–January for A. s. intermedius. Males call from emergent vegetation in ponds, dams and streams, in relatively high-density choruses of 4–20 individuals, between sunset and 03:00 (Backwell and Passmore 1991). The white eggs are deposited in longitudinally folded leaves of grasses, young Phragmites and herbaceous plants (particularly Commelina spp.), just below or above the water, and hatch 4–6 days later. Metamorphosis typically occurs within about six weeks (Backwell and Passmore 1991; Pickersgill 1996, 2000).
Predators include water snakes Lycodonomorphus spp., green snakes Philothamnus spp. and the Rhombic Night Adder Causus rhombeatus. Prey items include mosquitoes and other flies, spiders and moths (M.P. pers obs.).
During the atlas period, few records of this species were obtained (see table). Although not evaluated by Harrison et al. (2001), assessment for this publication placed it in the Vulnerable category on the basis of its extent of occurrence (<20 000 km2) and area of occupancy (<2000 km2), a severely fragmented range, a perceived continuing decline in area of occupancy, numbers of individuals and locations, and a decrease in the area and quality of its habitat.
The KwaZulu-Natal Nature Conservation Services has listed A. s. intermedius as a taxon of special significance because it is endemic to the province (Armstrong 2001).
Threats
The KwaZulu-Natal coast has several threatened frog species (Burger and Harrison 2002). This region has been extensively altered by urban and industrial developments, sugar cane plantations, timber plantations and infestations of alien vegetation, all of which have negatively affected biodiversity (Johnson 1987; Johnson and Raw 1987; Alexander 1990; Armstrong et al. 1998). These activities are the primary factors responsible for excessive loss and fragmentation of A. spinifrons habitat. Many of the surviving populations are small and localized and therefore more susceptible to catastrophic events that could result in local extinctions. Further threats to this species include pesticide pollution of breeding habitats, and trampling of habitat by livestock.
Recommended conservation actions
Historical localities should be revisited to establish the status of populations. In the course of such surveys, it is recommended that recordings of advertisement calls be collected along with specimens and tissue samples.
Armstrong (2001) noted that the species was inadequately conserved with only 2.8% of its predicted range falling within protected areas. A. spinifrons was predicted to occur in 12 KwaZulu-Natal protected areas, nine of which could potentially maintain viable populations, the largest being the uKhahlamba-Drakensberg Park (3864 ha). Although Armstrong’s (2001) assessment was based only on a subset of the species’ range, omitting the coastal populations, the species is clearly not adequately protected and it is therefore recommended that a series of known viable populations be evaluated with the aim of incorporating them into the network of protected areas. Protected areas in coastal KwaZulu-Natal, from which this species is known, include reserves at St. Lucia and Mtunzini. In Eastern Cape Province it is known from Hluleka and Silaka nature reserves.
Some general recommendations include habitat management, limiting factor management, monitoring, and public education. Future EIAs for urban, industrial, agricultural and forestry developments along the KwaZulu-Natal and Eastern Cape coast, should take cognisance of the conservation status of A. spinifrons and of potential impacts on its habitat, and should mitigate their plans accordingly.
Distribution
A. spinifrons is endemic to South Africa. The subspecies A. s. spinifrons occurs at low to intermediate altitudes from Cintsa Bay (3228CC) and Kei Road (3227DA) in Eastern Cape Province, to St. Lucia Village (2832AD) in KwaZulu-Natal. The subspecies A. s. intermedius occurs at altitudes above 1000 m in western KwaZulu-Natal, between Underberg (2929CD) and Hilton/Cedara (2930CB; Pickersgill 1996). In the course of the atlas survey, additional populations were discovered near Franklin (3029BC) and Umtata (3128DB). Populations in Eastern Cape and KwaZulu-Natal provinces, previously referred to A. knysnae by Poynton (1964), were assigned to A. s. spinifrons by Pickersgill (1996).
Unlike most frog species sharing its breeding habitat, A. spinifrons has a rather quiet call. This may be a factor contributing to the paucity of atlas records and the patchy distribution reflected on the map.
Distribution of Afrixalus spinifrons. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Natal Spiny Reed Frog, Spinose Dwarf Leaf-folding Frog, Natalse Blaarvouendepadda (Afrikaans)
Recommended citation format for this species text:
Pickersgill M, Burger M, Bishop PJ, Tippett RM. Natal Leaf-folding Frog Afrixalus spinifrons. BDI, Cape Town. Available online at http://thebdi.org/2022/01/21/natal-leaf-folding-frog-afrixalus-spinifrons/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Pickersgill M, Burger M, Bishop PJ 2004 Afrixalus spinifrons Natal Leaf-folding Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Alison Sharp) in FrogMAPhere.
Find the Golden Leaf-folding Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Hyperoliidae
GOLDEN LEAF-FOLDING FROG – Afrixalus aureus
Pickersgill, 1984
Habitat
A. aureus is a savanna species which inhabits Coastal Bushveld-Grassland, a mosaic of vegetation types found from sea level to an altitude of 300 m along the coast of northern KwaZulu-Natal, as well as various other bushveld vegetation types, such as Mixed Lowveld Bushveld, at altitudes of 200–300 m, east of the eastern escarpment (Jacobsen 1989). Along the coast, it seems to prefer drier habitats than those occupied by A. delicatus and A.spinifrons and it does not usually utilize the same breeding sites as the latter species (Pickersgill 1984; M.P. pers. obs.).
Breeding takes place in seasonal or semi-permanent water bodies containing emergent vegetation such as Polygonum pulchrum, Ludwigia stolonifera and Cyperuspapyrus (Backwell 1991). At most breeding sites the water is fairly shallow, rarely exceeding 50 cm in depth.
Behaviour
Nothing is known of the non-breeding behaviour of this species.
During the breeding season (November–February), adults can be found close to breeding sites at the margins of pans and grassy pools.
Afrixalus aureus – Manyeleti Game Reserve, Mpumalanga Photo by Vaughan Jessnitz
The frogs may sit on emergent vegetation, usually in leaf axils or on broad-leaved shrubs, fully exposed to the sun, in a head-down position. At dusk, males take up positions a few centimetres above the water, where they begin to call, continuing until about 04:00. High-density choruses develop; these contain a relatively high proportion (50%) of satellite males and are characterized by a high level of aggression (Backwell 1991). Within the choruses, males do not form small groups, as in A. spinifrons and A. delicatus.
Amplexus is axillary, and clutches of about 50 creamy white eggs are enveloped in vertically folded leaves (often of Polygonum pulchrum or Ludwigia stolonifera) or blades of grass, 4–15 cm above the surface of the water (Jacobsen 1989; Backwell 1991; Channing 2001).
Afrixalus aureus – Pair during amplexus Near Hoedspruit, Limpopo Photo by Luke Verburgt
Status and Conservation
A. aureus is not considered to be threatened. Its habitat is used for stock and game farming, as well as ecotourism. It has been observed to breed in cattle waterholes, where vegetation is relatively sparse. It occurs in a number of provincial nature reserves, such as False Bay, Hluhluwe, Mkuze, Umfolozi, Nyalazi and Tembe, and in Kruger National Park.
Distribution
This species ranges northward into southern Mozambique (Pickersgill 1984; Poynton and Broadley 1987). Within the atlas region, A. aureusisa locally abundant species thatoccurs from the northeastern parts of the Limpopo Province, southward through eastern Mpumalanga, central and eastern Swaziland, to northern KwaZulu-Natal.
It is usually abundant at its breeding sites and easy to identify by its call. Although A. aureus and A. delicatus are morphologically similar, their calls are quite distinct and they usually use different breeding sites. The atlas data for A. aureus are reasonably complete and reliable.
Distribution of Afrixalus aureus. Taken from the FrogMAP database, April 2022
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Golden Spiny Reed Frog, Golden Dwarf Leaf-folding Frog, Goue Stekel-rietpaddatjie (Afrikaans), Goue Blaarvouendepadda (Afrikaans)
Recommended citation format for this species text:
Pickersgill M, Bishop PJ, Tippett RM. Golden Leaf-folding Frog Afrixalus aureus. BDI, Cape Town. Available online at http://thebdi.org/2022/01/21/golden-leaf-folding-frog-afrixalus-aureus/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Pickersgill M, Bishop PJ 2004 Afrixalus aureus Golden Leaf-folding Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Allison Sharp) in FrogMAPhere.
Find the Mottled Shovel-nosed Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Hemisotidae
MOTTLED SHOVEL-NOSED FROG –Hemisus marmoratus
Peters, 1854
Identification
Hemisus marmoratus – Near Hluhluwe, KwaZulu-Natal Photo by Ryan Tippett
Habitat
This species thrives in semi-arid environments and is well-adapted to breeding in shallow, temporary water bodies. In the atlas region it inhabits a variety of bushveld vegetation types in the Savanna Biome.
Breeding habitat includes pans, waterholes or artificial impoundments, as well as the isolated pools that form in riverbeds as water levels drop. The substrate usually consists of fine mud or clay, but burrows have been observed in coarser sandy sediments too.
In West Africa, H. marmoratus is found mainly in savanna habitats but also occurs in forest, and tadpoles have been encountered in a wide range of water bodies, except major rivers (Rödel 2000).
Behaviour
These burrowing frogs spend the dry season in a torpid state, underground. They begin to call as soon as the first spring rains have soaked the ground, sometimes even before standing water has accumulated at the breeding site. At this time they construct extensive, shallow, tortuous tunnels, in low muddy areas that are likely to fill with subsequent rains, or close to the edges of pools that have already formed. The tunnels form conspicuous low ridges on the surface, often intersect, and sometimes terminate in larger, rounded chambers. Males usually leave the tunnels and call from the surface, but in the absence of ground cover they may call from within the tunnels or chambers (pers. obs). Food includes ants, termites and earthworms (Rödel 2000; Channing 2001).
Hemisus marmoratus – Kruger National Park, Mpumalanga Photo by Cornelia Rautenbach
Choruses at Hans Merensky Nature Reserve typically comprise groups of 5–8 males that participate in discrete bouts of calling, alternating with periods of silence (pers. obs.). One male usually initiates calling, followed by the other members of the group. Calling males may be as little as 30 cm apart.
Amplexus is inguinal. Once in amplexus, the female selects a suitable oviposition site and disappears beneath the surface, male in tow, to excavate a nest. Nests may be constructed in low-lying areas that are subsequently flooded after rain (Kaminsky et al. 1999), or in more elevated positions away from the water’s edge. For example, nests have been found 15 cm to 8 m from the water’s edge in South Africa (pers. obs.; Jacobsen 1989), while in Comoé National Park in Ivory Coast, nests have been found up to 100 m from the nearest pool (Rödel 2000).
A chamber is constructed in which a spherical mass of 88–242 eggs are laid (Rödel 2000). Sterile jelly capsules are laid on top of the fertile eggs and the entire mass is bound together by a fibrous substance that prevents the egg mass from being flattened by the female, who sits on top of the mass until the eggs hatch. After hatching, the developing tadpoles cling to the body of their mother, who actively defends them against intruders (Rödel et al. 1995).
In dry weather, development is arrested and the tadpoles can remain in the chambers for as long as two months in anticipation of the next rains (Kaminsky et al. 1999). If rising water floods the nest, the tadpoles leave it (at any stage of development) and enter the water to feed. If the nest is not flooded, the female provides an escape route from the nest to the water by constructing a surface slide down which the tadpoles wriggle to the water (Rödel et al. 1995; Kaminsky et al. 1999). If this is not possible, the female may carry her tadpoles to the water while they adhere to her body.
Since the tadpoles of H. marmoratus often begin their development before the breeding site contains water, they have an advantage over tadpoles of species that lay their eggs only after heavy rain in that, once the site is flooded, Hemisus tadpoles take less time to reach metamorphosis and are exposed to predators for a shorter period of time. These adaptations are advantageous in an environment where rainfall is unpredictable and highly variable.
The species does not appear to be at risk, as much of its habitat is used for game and cattle farming and is relatively undisturbed. It occurs in a number of provincial nature reserves and national parks.
H. marmoratus is a wide-ranging inhabitant of the savannas of sub-Saharan Africa, from Senegal in the west, to Ethiopia and Somalia in the east, and southward to Angola, eastern Namibia (Caprivi), Botswana, Zimbabwe and Mozambique (Poynton and Broadley 1985a).
In the atlas region the species is distributed from Derdepoort (2426DA) in the far western corner of Limpopo Province, eastward across the dry northern and central parts of the province into the lowveld, and south through eastern Mpumalanga and central and eastern Swaziland to northern KwaZulu-Natal, reaching Mfolozi (2831BD) and Ntambanana district (2831DB) in the south. In 1999, it was recorded in Gauteng Province in a borrow pit at Burkea Park near Pretoria, but this outlying record may represent an accidental translocation.
A morphologically similar species, H. guineensisbroadleyi, apparently reaches the southern limits of its range north of the Soutpansberg in Limpopo Province, where it is known from two historical records. This species can be distinguished from H. marmoratus by differences in its advertisement call (see species account for H. g. broadleyi).
The atlas data for H. marmoratus are reliable but incomplete.
Distribution of Hemisus marmoratus. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Marmergraafneuspadda (Afrikaans)
Recommended citation format for this species text:
Minter LR, Tippett RM. Mottled Shovel-nosed Frog Hemisus marmoratus. BDI, Cape Town. Available online at http://thebdi.org/2022/01/21/mottled-shovel-nosed-frog-hemisus-marmoratus/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Minter LR 2004 Hemisus Marmoratus Mottled Shovel-nosed Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Table Mountain Ghost Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Heleophrynidae
Identification
Table Mountain Ghost Frog males reach 50mm and females attain 63mm in snout-vent length. They resemble other Heleophryne speciesin their body shape and posture, long limbs, and large spatulate, adhesive pads at the tips of their fingers and toes.
The dorsum is green with reddish-brown to purple mottling, while the ventrum is granular and generally white with pink shading.
Habitat
The Table Mountain Ghost Frog inhabits clear perennial streams situated in moist, forested ravines, gorges and valleys, or in fynbos on the more exposed slopes and shallow valleys, especially in the upper catchment areas. The vegetation types in these habitats are Afromontane Forest and Mountain Fynbos, respectively. The streams are rocky and fast-flowing for most of the year, forming waterfalls interspersed with slower-flowing sections and pools. (Boycott and De Villiers 1986).
Following metamorphosis, froglets move out of the streams to damp boulders, moss-covered rock faces and crevices close to the streams. Non-breeding adults have been found in damp, sheltered habitat well away from the streams, including caves (Gow 1963; Boycott and De Villiers 1986).
Behaviour
The Table Mountain Ghost Frog is cryptic and elusive and occurs in relatively low numbers. Its typical habitat is often inaccessible and consequently, there is limited information on breeding activity and general life history.
Breeding takes place in spring and summer, once the winter rains have ended and there is a reduction in stream flow (Boycott and De Villiers 1986). Skin asperities, a secondary sexual character, have been noticed in late August and are well developed by November and December. Gravid female frogs have been collected October–December, and a call was recorded in December 1978 (see sonagram in Passmore and Carruthers 1995). The eggs and oviposition site have not been recorded.
The distinctive tadpoles attain a length of 40–50 mm and are adapted to torrents. They have large, sucker-like oral discs which enable them to cling to rocky substrates in fast-flowing streams and to climb slippery, vertical rock faces. The tadpoles feed on algae covering the rock surfaces, and their feeding trails can be seen on rocks in quieter pools. During the day they are usually found attached to the underside of submerged rocks. H. rosei tadpoles take >12 months to complete metamorphosis, and are ready to leave the water by April or May.
Table Mountain Ghost FrogHeleophryne rosei tadpoles Skeleton Gorge, Western Cape Photo by Pieter la Grange
Status and Conservation
Status
The Table Mountain Ghost Frog was previously listed as Rare (McLachlan 1978) and Endangered (Branch 1988). The species is now classified as Critically Endangered, owing to its small extent of occurrence and area of occupancy (regarded as a single location) and a projected decline in the area of occupancy, habitat quality and number of mature individuals (Harrison et al. 2001; this publication). The species is legally protected by Nature Conservation Ordinance 19 of 1974. At present H. rosei is not listed by CITES.
Although H. rosei has suffered some habitat loss, this does not appear to have led to habitat fragmentation. However, the construction of two large reservoirs in Disa Stream and three in Original Disa Stream, between 1890 and 1907, must have obliterated a significant proportion of prime H. rosei habitat. The disappearance of the tadpoles of this species from Platteklip Gorge by 1980 may be attributed to climatic change, erosion and the spread of alien vegetation, while water abstraction and stands of alien trees are probably the main cause of its absence from Cecilia Ravine since 1997. It is estimated that the number of tadpoles in Skeleton Gorge has decreased by 50% since 1980. In total, 20% of the habitat of this species is estimated to have been lost since 1980.
Since 1997, systematic tadpole counts have been carried out in all streams at the end of summer when stream levels are at their lowest. At each monitoring site, tadpoles are counted over a 30-min period during the day. Between 1997 and 2001, <20 tadpoles were counted at each site, and on a number of occasions, <10 were seen (unpubl. data). This indicates that the species survives at low population densities and suggests that there has been a population decline since the earlier surveys were conducted.
Fortunately, all habitat of H. rosei is now protected within the recently proclaimed Cape Peninsula National Park and the undeveloped upper reaches of Kirstenbosch National Botanical Garden. In addition, the extensive clearing of pine trees and other alien vegetation on Table Mountain since 1980 has helped to improve the quality of H. rosei habitat in places.
Threats
Although H. rosei occurs within a protected area, its survival is not assured. For example, in other parts of the world, amphibian species have disappeared from protected mountainous regions as a result of the introduction of predatory fish, disease or environmental deterioration (Beebee 1997; Berger et al. 1998; Longcore et al. 1999). The fact that Table Mountain is situated next to a large city and is extensively utilized by human visitors (including foreign tourists), increases the possibility of introduction of disease.
Since metamorphosis in H. rosei takes >12 months, an uninterrupted flow of water is essential to the survival of the tadpoles. Consequently, global warming poses a serious threat if reduced rainfall and prolonged droughts lead to a loss of perennial streams. At present, most of the streams are reduced to a trickle by the end of summer, and few tadpoles are present. Similarly, large reservoirs in Disa Stream and Original Disa Stream represent a further potential threat to the water resources for tadpole populations. Elsewhere, water abstraction takes place in Cecilia Ravine and in the lower reaches of Window Gorge. It is essential to maintain the seasonal flow below dams throughout the year if tadpole populations are to survive in these parts of the streams (Boycott and De Villiers 1986; Boycott 1988c; Harrison et al. 2001). Invasive alien vegetation and plantations of exotic trees have also reduced runoff and stream flow. Furthermore, the presence of alien deciduous trees alongside streams (e.g. poplars) leads to the severe clogging of tadpole habitat with leaf-litter (Boycott and De Villiers 1986).
The effect of erosion on H. rosei habitat is difficult to determine, but increased siltation is believed to have caused the deterioration or loss of tadpole habitat in places. The erosion problem on Table Mountain is caused by alien vegetation and the removal of dense stands of introduced species, trampling by human pedestrian traffic, overgrazing and trampling by the invasive Himalayan Tahr, and too frequent fires (Moll and Campbell 1976).
Recommended conservation actions
The distribution and relative abundance of H. rosei is monitored by the Western Cape Nature Conservation Board as part of a threatened species monitoring programme (De Villiers 1997a), while the Cape Peninsula National Park and Kirstenbosch National Botanical Garden are responsible for the protection and management of the habitat of the species. In managing the habitat, priority is given to erosion control, eradication of alien vegetation and the regulation and maintenance of streamflow from the various reservoirs.
The monitoring of tadpole populations should be intensified to obtain a more accurate estimate of population size and its fluctuation from year to year. This will produce new information on the reproductive biology and phenology of the species.
There is potential for decline at the hands of unscrupulous collectors. Additional protection against illegal trade could be afforded by listing H. rosei in one of the CITES appendices.
Boycott and De Villiers (1986) suggested that the translocation of tadpoles to other streams on the mountain be considered. However, this is not recommended at present in view of the low numbers of tadpoles in known localities and the absence of suitable perennial streams in which the survival of the tadpoles could be guaranteed. A captive breeding programme for H. rosei is not recommended. The stocking of reservoirs and streams with predatory alien fish should be strictly prohibited for obvious reasons.
Distribution
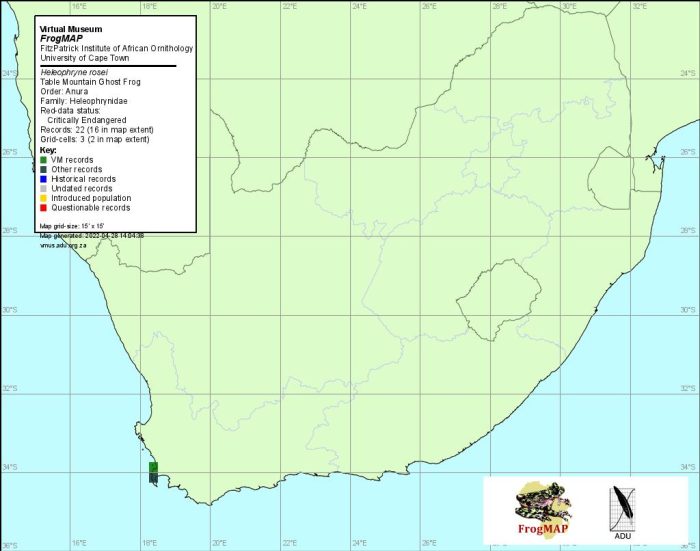
The Table Mountain Ghost Frog occurs in the winter-rainfall region of the Western Cape and is endemic to Table Mountain on the Cape Peninsula (3318CD). It occupies an area of <10 km2 at an altitude of 240–1060 m. With the exception of a section of stream on top of the mountain which drains the western slopes, and one that drains the northern slope (historical record), H. rosei occurs mostly in streams draining the wetter southern and eastern slopes that receive annual rainfall of 1600–2000 mm.
Distribution of Heleophryne rosei. Taken from the FrogMAP database, April 2022.
During the 50-year period following its description, this rare and elusive frog was recorded from only a few scattered localities: Skeleton Gorge above Newlands, Wynberg Caves, Platteklip Gorge, the top of Window Gorge, upper Disa Stream, and Echo Valley near Grotto Caves (Hewitt 1925; Du Toit 1934; Gow 1963; Poynton 1964; records of the Western Cape Nature Conservation Board).
In 1980, a systematic survey recorded tadpoles in six streams (Boycott and De Villiers 1986). On the south-facing slopes they were found in Disa Stream (upper catchment section and below the reservoirs) and Original Disa Stream. On the east-facing slopes they were recorded from Window Gorge, Skeleton Gorge, Nursery Ravine and Cecilia Ravine. There were no signs of H.rosei on the northern and western slopes, including Platteklip Gorge, a previously recorded locality. During this survey, one adult and a few froglets were found in addition to tadpoles.
The most recent survey formed part of a threatened amphibian monitoring programme (De Villiers 1997a) and took place during 1997–1999. Tadpoles were found at all localities recorded during the 1980 survey except Cecilia Ravine. A new locality was discovered on top of the mountain in the upper section of a stream which drains into Kasteelspoort, in the Valley of the Red Gods. This represents the only confirmed record of the species in a stream that drains the west-facing slopes (unpubl. data).
In summary, H. rosei tadpoles have been recorded in a total of eight streams but the species is currently known to breed in only six. The atlas distribution data are reliable.
Further Resources
The use of photographs by Megan Loftie Eaton and Pieter la Grange is acknowledged.
Other Common Names: Tafelberg-spookpadda (Afrikaans).
Recommended citation format: de Villiers, AL; Tippett, RM. (2025). Table Mountain Ghost Frog Heleophryne rosei. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2022/01/21/table-mountain-ghost-frog-heleophryne-rosei/
This species text has been updated and expanded from the text in the 2004 frog atlas: de Villiers, AL. (2004). Table Mountain Ghost Frog Heleophryne rosei. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
There are currently no photographs or maps available for this species.
Find Hewitt’s Ghost Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Heleophrynidae
Identification
Hewitt’s Ghost Frog is a medium-sized species attaining a snout-vent length of 50mm in females and 47mm in males. This species has the long legs, spatulate digits, and squat posture characteristic of Heleophryne. The upperparts are light brown to olive-brown with large, rounded to irregular dark blotches, that have thin, pale margins. The blotches form dark transverse bands on the legs.
Habitat
Hewitt’s Ghost Frog inhabits the upper reaches of clear, swift-flowing, perennial mountain streams with rocky beds, in an area with annual rainfall of c.1000 mm. Adults and tadpoles are found beneath submerged and partly-submerged rocks in these streams, and occasionally at the edge of small waterfalls and cascades (Boycott and Branch 1988). It is assumed that the adults leave the streams during the non-breeding season to forage in surrounding terrestrial habitat, as is the case in other Heleophryne species.
The Elandsberg range lies within the Fynbos Biome, and H. hewitti occurs in Grassy Fynbos on the southern, relatively mesic slopes. However, there are only remnant patches of fynbos because most of the natural vegetation has been replaced by plantations of exotic trees that extend virtually onto the stream banks. Thus little non-breeding habitat is available to this species.
Behaviour
Nothing is known of the non-breeding behaviour of H.hewitti. Calling males have been heard October–November, indicating a similar breeding period to that of H. purcelli and H. orientalis. Males call at night and during the day under large, flat, partly submerged rocks in shallow, fast-flowing sections of streams. These rocks usually rest on top of smaller rocks, and the calling males take refuge in the spaces between the latter. Males have also been found calling from the sides of rocky gullies through which the stream flows, taking up positions directly above and within centimetres of the rushing water.
Gravid and spent females and eight egg batches were found in October (Boycott 1988). The eggs were large and yellow and numbered 93–150 per clutch. They were laid under submerged or partly submerged rocks of variable size, usually in relatively quiet backwaters where stream flow was not rapid and water depth was 20–60 cm. However, one batch was attached to the underside of a rock in a shallow, fast-flowing stream. As with all species of ghost frog, the tadpoles of H. hewitti are more easily located than the adults and, in some instances, distribution records for the species have been based solely on tadpoles. Tadpoles and adults are preyed upon by the common Brown Water Snake Lycodonomorphus rufulus (pers. obs.).
Status and Conservation
Status
H. hewitti was previously listed as Endangered (Branch 1988). Its present status is Critically Endangered, based primarily on an extent of occurrence <100 km², as well as substantial loss, fragmentation and degradation of its habitat, and projected decline in its area of occupancy and the absence of protection within a nature reserve (Harrison et al. 2001; this publication).
Heleophryne hewitti is a protected species under the Cape Nature Conservation Ordinance (Ordinance 19 of 1974, as amended) and the SAFCOL (South African Forestry Company Limited) Environmental Policy, according to the standards of the Forest Stewardship Council (FSC) (Harrison et al. 2001).
Threats
H. hewitti appears to have one of the most restricted distribution ranges of any southern African amphibian, and that in an area dominated by alien pine plantations. The species is threatened by habitat loss, fragmentation and degradation as a result of over-utilization of its habitat for forestry, frequent and sometimes devastating forest fires and floods. In the 1980s, mountain fires devastated the whole region and during subsequent clean-up operations, many burnt trees were felled and left in the watercourses. Thereafter, heavy rains washed much of the felled timber and debris into the streams, blocking the flow and causing siltation of virtually every stretch of Heleophryne habitat, including the type locality. Other possible threats to the species include the construction of dams, and the introduction of alien fish species (Boycott and Branch 1988; Harrison et al. 2001).
Recommended conservation actions
Priority needs to be given to the conservation and management of remaining H. hewitti breeding and non-breeding habitats. The absence of protection for H.hewitti within a provincial or private nature reserve is a serious concern. Harrison et al. (2001) recommended monitoring of known populations, an intensive search for new populations, and an investigation of the taxonomic relationship between H. hewitti and other Heleophryne populations in the vicinity. They also suggested that a population and habitat viability assessment (PHVA) be carried out and that the public be informed of the plight of this species.
Over the last two decades the habitat of H. hewitti has deteriorated alarmingly. This may be attributed solely to mismanagement of, and blatant disregard for, the habitat of an endangered species. H. hewitti was classified as an endangered species in 1988, shortly after its discovery. During a June 2002 visit to the Geelhoutboom River type locality and Martins River, the habitat was found in a shocking condition. Both localities were heavily infested with alien pine and wattle trees. In particular, the type locality was sedimented to such a degree that pine trees were growing in the riverbed. On account of the sedimentation there has been a considerable reduction in the breeding habitat where riffles, rapids and rocky pools have been buried under sand. High levels of algal growth were evident, probably as a result of the elimination of invertebrate fauna and the loss of direct sunlight due to trees growing in and over the river.
To recover some of the former breeding habitat, it is imperative that action be taken immediately. All alien trees (pine and wattle) must be cut down and removed from the riverbed and the riverbanks. These should be cut and removed manually from the site and not left to lie and rot in the rivers or on the banks. In order to expose the bedrock and other rocky sections of the river, removal of sand from the riverbed should be considered.
Distribution
Hewitt’s Ghost frog is endemic to the Eastern Cape Province of South Africa, where it is confined to the Elandsberg Range. It is restricted to four rivers on the Elandsberg Range: the Geelhoutboom, Martin’s, Klein and Diepkloof rivers. Only the Martin’s and Klein rivers have perennial tributaries, and the sources of the four rivers extend over a distance of just 10 km. The greatest distance separating any two of these rivers is 3.5 km. The recorded altitudinal range is 400–550 m (Boycott and Branch 1988). Historically the species is only known from three quarter-degree grid cells, and no further grid cells were added during the atlas period.
The relationship between H. hewitti and presently unidentified Heleophryne populations in the Kammanassie, Kouga and Baviaanskloof mountains, requires investigation (Boycott and Branch 1988; Branch and Bauer 1995; Harrison et al. 2001). Tadpoles recently collected from the southeastern slopes of Cockscomb (3324DB) in the Grootwinterhoek Mountains were tentatively identified as H. hewitti, but await verification (M. Cunningham pers. comm.)
Other Common Names: Hewitt se Spookpadda (Afrikaans).
Recommended citation format: Boycott, RC; Tippett, RM. (2025). Hewitt’s Ghost Frog Heleophryne hewitti. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2022/01/20/hewitts-ghost-frog-heleophryne-hewitti/
This species text has been updated and expanded from the text in the 2004 frog atlas: Boycott, RC. (2004). Hewitt’s Ghost Frog Heleophryne hewitti. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
View the above photo record (by Arnold van der Westhuizen) in FrogMAPhere.
Find the Sand Toad in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
SAND TOAD – Vandijkophrynus angusticeps
Smith, 1848
Habitat
V. angusticeps generally inhabit sandy soils in areas of high winter rainfall in the Fynbos Biome. Breeding occurs in shallow temporary pools in seasonally flooded fynbos or renosterveld vegetation types. In some areas, breeding sites may be in ploughed and planted fields, such as in the Swartland around Vredenburg (3217DD), and near Moorreesburg (3318AB) where this species co-occurs with Cacosternum capense on clay soils. V. angusticeps is not commonly seen outside the breeding season, although in montane sites individuals have been found sheltering under rocks (Branch 1990). The type series was “procured at different seasons of the year . . . among decaying leaves in districts coated with underwood” (Smith 1848).
Behaviour
Breeding occurs in winter, May–September, after heavy rains have saturated the soil and created pools that last for many weeks. On nights after such rains large numbers of V. angusticeps may emerge from their refuges and move to breeding sites. Numerous individuals may be encountered on roads at this time, and may move considerable distances to reach breeding sites.
In comparison with the frenzied cacophony of most toad choruses, breeding aggregations of V. angusticeps are a subtle affair. Calling males are often sparsely distributed and, as with several species in the Angusticeps Division, such as V. amatolicus, calls are weak and intermittent. Males call from exposed sites near the water’s edge. Rose (1929) recorded 650–850 eggs in a clutch, relatively few for a toad, while Channing (2001) set the upper limit at 3000. Blair (1972) noted that species in the Angusticeps Division have exceptionally large testes. These characteristics suggest strong competition among males, and that V. angusticeps uses other modes of communication to locate and choose mates. A study of the mating system of this species would provide a very interesting comparison with other southern African toads.
Rose (1962) recorded insects and small snails as food items.
Status and Conservation
The habitat of V. angusticeps is fragmented among large areas of coastal sand flats and montane isolates, and has declined in abundance due to development of the coastal flats and drainage of seasonally inundated wetlands. It has been estimated that these threats have resulted in a greater than 50% reduction in the abundance of V. angusticeps over the past century (Harrison et al. 2001). In the past 20 years, the species has virtually disappeared from sites where it was formerly abundant, such as Rondebosch Common in Cape Town (Rose 1929) and the Jan Marais Municipal Nature Reserve in Stellenbosch (E. van Dijk pers. comm.).
Nevertheless, this species still has a relatively large extent of occurrence and area of occupancy away from urban centres, and is present in large protected areas such as Kogelberg Biosphere Reserve (3419AC), De Hoop Nature Reserve (3420AD) and Cape Peninsula National Park (3418AD). Hence, V. angusticeps was assigned to the category Least Concern (Harrison et al. 2001; this publication). Its conservation status will require reassessment once the taxonomic status of inland and montane populations have been determined.
Distribution
V. angusticeps is endemic to the Fynbos Biome, within which it is widespread, extending from near Humansdorp (3424BA) in the east, and along the coastal flats and Cape fold mountains to Nieuwoudtville (3119AC) in the west. Most quarter-degree grid cell records (almost 95%) are located in the Western Cape Province. Within this area, the species occurs in two very distinct environments, namely coastal flats and rocky montane areas.
Most records are from the sandy coastal flats between Witsand (3420BD), at the mouth of the Breede River, and Vredenburg (3217DD) near the Berg River mouth. V. angusticeps is rarely encountered in the eastern part of its distribution, probably partly due to the limited area of coastal sand flats along the Garden Route between Mossel Bay and the Tsitsikamma River mouth. More surveys are needed, however, in the extensive coastal sand flats of the Stilbaai area, between the Breede and Gouritz rivers (3421AC–3421BD).
Montane records of this species are from scattered sites high in the Cape fold mountains, such as around 1500 m in the Swartberg Mountains (3322AC; Branch 1990; 3321BD new atlas data) and near the summit of Matroosberg (3319BD; Branch 1990). Smith (1848) described the type locality as “the interior of Southern Africa” and “in localities far apart”, which is barely more accurate than specific.
V. angusticeps is part of an extremely closely related and morphologically confusing group of toads endemic to southern Africa that also includes V. gariepensisgariepensis, V. nubicolus, V. inyangae, V. robinsoni and V. amatolicus (Cunningham and Cherry 2000). Most of these taxa are geographically variable and at some sites it is difficult to assign individuals to a particular species.
Beyond the first range of Cape fold mountains, in the Little Karoo, Bokkeveld and ranges bordering the Great Karoo, the distribution of V. angusticeps overlaps that of V. gariepensis, although these species are rarely found in true sympatry and some shared locations may be based on incorrect identifications. Boycott (1988d) commented that the small toads from mountain fynbos in the Kammanassieberg (3322DB) were similar to V. amatolicus, although these are well outside the range of that species and should probably be considered a montane variant of V. angusticeps.
In the west, V. angusticeps is distributed across the Olifants River, overlapping the ranges of both V.gariepensis and V. robinsoni in the vicinity of Vanrhynsdorp. In this area, V. angusticeps appears to be displaced from the lowlands and is only found on the Bokkeveld escarpment and Gifberg/Matsikamma Mountain at the northwestern limit of the Cape fold mountains (3118DC; new atlas data). Poynton (1964) suggested that specimens collected in Namaqualand, between latitudes 30–31°S, are intergrades between taxa. Clearly more research is needed on morphology, colouration and genetic variation within this group, and this will require extensive and systematic sampling across the entire distribution of the Angusticeps Division, including montane populations and contact zones between taxa. Typical individuals of V. angusticeps are distinguished from other species in the Angusticeps Division by their unspotted white ventrum and bright yellow flush over the dorsal surfaces of their feet. However, these characters may be absent in certain areas, making identification difficult.
The atlas data are reasonably reliable but incomplete.
Distribution of Vandijkophrynus angusticeps. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Cape Sand Toad,Sand Toad, Yellow-footed Toad (Alternative English Names); Sandskurwepadda (Afrikaans)
Recommended citation format for this species text:
Cunningham M, Tippett RM. Sand Toad Vandijkophrynus angusticeps. BDI, Cape Town. Available online at http://thebdi.org/2022/01/20/sand-toad-vandijkophrynus-angusticeps/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Cunningham M 2004 Vandijkophrynus angusticeps Sand Toad. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.







































{kind=link}
{kind=link}
{kind=link}
{kind=link}