If you enjoy wildlife documentaries or scientific articles, you’ve probably seen photos or video taken by “camera traps.” Remote cameras have been used for years by scientists studying or documenting wildlife numbers and behaviour in sometimes difficult or inaccessible terrain. A camera trap is just like an ordinary digital camera except that you don’t have to press a button for a photo to be take, instead the camera is triggered by movement or heat from animals. Camera traps are super handy tools for mapping mammals, they are like little spies in the bush!
Camera trapping has proved to be a very effective way of finding out which elusive and, especially, nocturnal animals are in an area. It’s also an effective way to find out how animals are utilising an area.
Porcupines coming for a drink with a Kudu strolling on by
There is an Animal Demography Unit project for which camera traps play an absolutely crucial role and that is MammalMAP. MammalMAP is the Atlas of African Mammals. The aim of MammalMAP is to update the distribution records for all of Africa’s wild mammals — the small ones, the big ones, the dry ones and the wet ones.
“Surely we know the distributions of Africa’s mammals? These are flagships species for tourism in Africa.” Sadly, the answer is “no” — the distributions are changing due to habitat destruction and climate change. Developing these 21st century distribution maps is filling a critical gap in conservation needs. To effectively manage and conserve wildlife we need to know where they are and we need to understand why they are there. But the reality is that across Africa, our knowledge of the whereabouts of many mammals is, at best – outdated, and, at worst – based on unverified anecdotes. Filling this crucial gap in our knowledge is the main aim of MammalMAP. If you are keen to help us map Africa’s wonderful mammals then setting up a few camera traps is a fun and effective way to do so.
A few tips on setting up camera traps:
It is very important to pick the right site for your camera trap. It helps to be quite sure that an animal will pass by the camera at some stage
Well-used game paths, hiking trails, quiet jeep tracks, dry watercourses and the bottom of ravines are all good places to set up your camera traps
For close shots, like on game paths, at watering holes or food sources set the camera up at no higher than waist height (if you are focusing on smaller animals it is better to set the camera up at a lower position)
If you are in an area where elephants, hyenas or large predators occur, make sure that your hands are free from any unusual or attractive odours e.g. food, perfumes etc. as this might tempt these animals to inspect the source of the odour — and they may just have a pull or a bite at the camera trap to see if its to their liking
Another important factor to keep in mind is the direction of the rising and setting sun. Occasionally when a subject triggers the camera when it is pointed towards the sun as the sun is rising or setting (typically heavy activity hours) this can lead to overexposed or ‘washed-out’ pictures
A leopard caught on camera mid hunt!
You can upload your camera trap photos to MammalMAP at http://vmus.adu.org.za — have fun!
The Hantam is the general area north of Calvinia in the Northern Cape province of South Africa. It’s an arid area, and poorly covered in biodiversity surveys. Citizen scientist Salome Willemse arranged a BioBash for the area, the “Calvinia BioBash”, and found accommodation in a farm house called “’n Handvol Gruis”. The expression, “’n handvol gruis”, literally, “a handful of grit”, comes from a poem by C Louis Leipoldt:
The last verse of “‘n handvol gruis” by C. Louis Leipoldt
For this poet, C. Louis Leipoldt, this area, the Hantam-wyk, was one of the most beautiful places he had experienced. I searched for an English version of this poem which captures the rich mood, but Wikipedia says that Leipoldt’s poetry doesn’t translate easily. In the last line, “arm” means “poor”, “eergister” is “the day before yesterday”, “en” is “and”, “nou” is “now”, and “skatryk” is “treasure rich”. Literally: “Poor the day before yesterday, and now treasure rich.”
Not everyone sees beauty here, but Leipoldt did!
The last line of the poem is about an emotional transformation from poverty to riches. The Calvinia BioBash aimed to make the same transformation, but in a somewhat more practical way. We aimed to transform the biodiversity database of the Hantam from poverty to riches. We certainly didn’t make it “skatryk”, but we enhanced the quality of the data dramatically.
Awesome accommodation for the BioBash. This farmhouse is called ‘n Handvol Gruis, perpetuating the memory of the poet. If you are looking for accommodation in this area, this is a great place to stay.
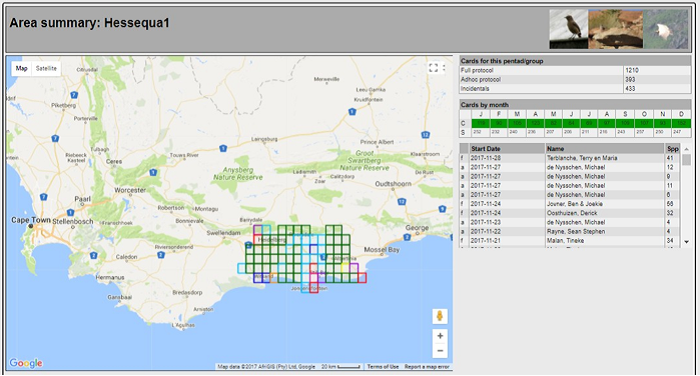
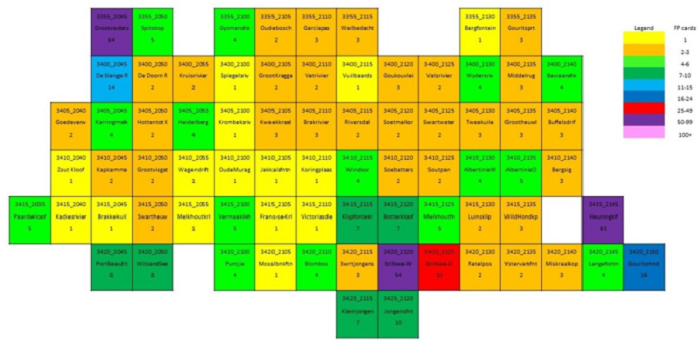
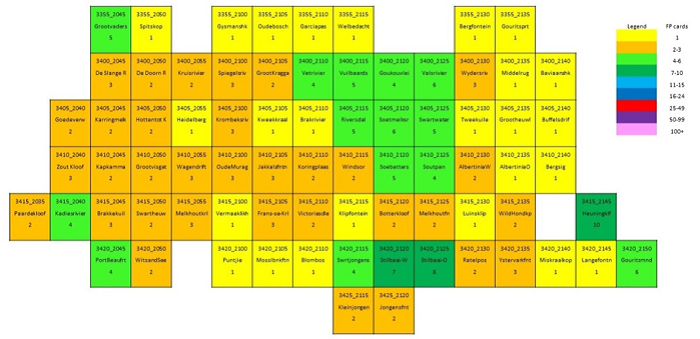
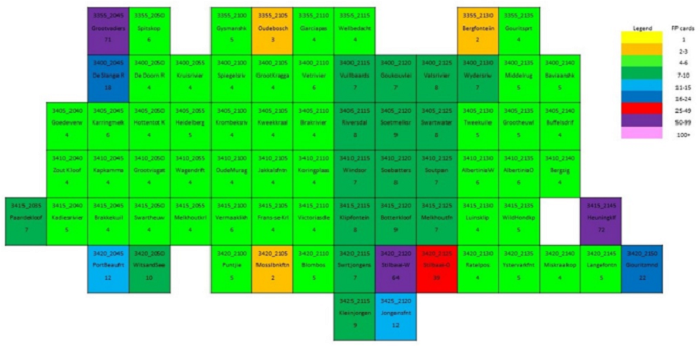
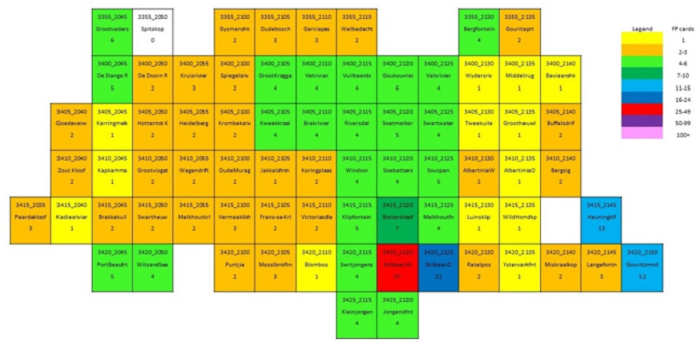
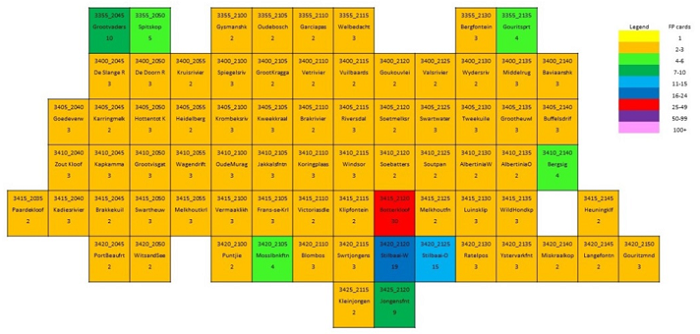
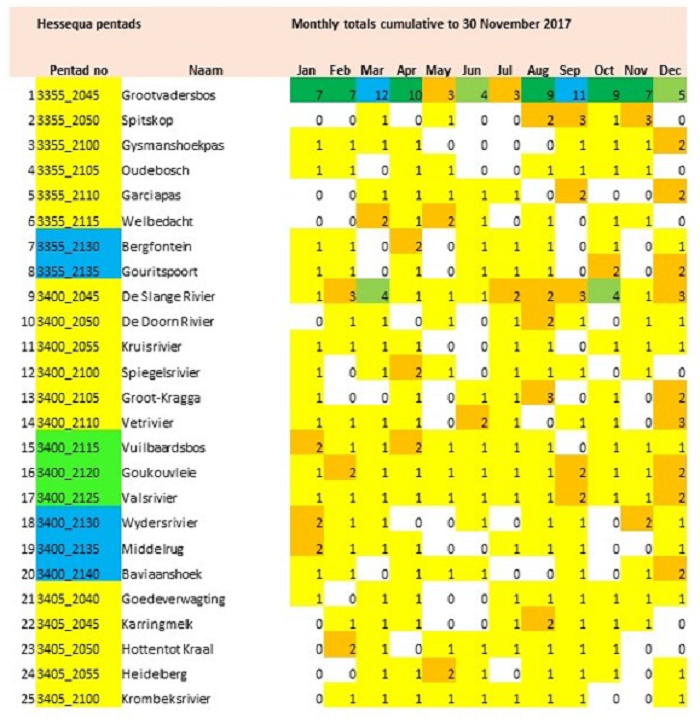
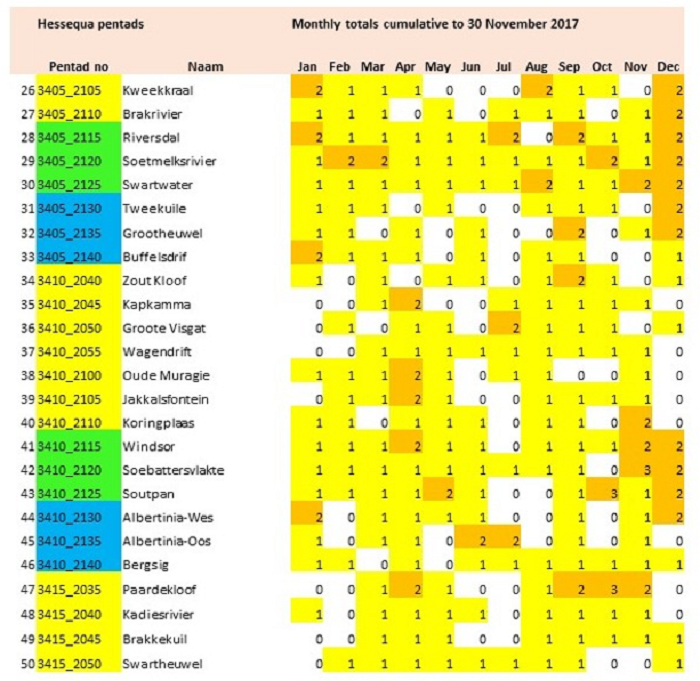
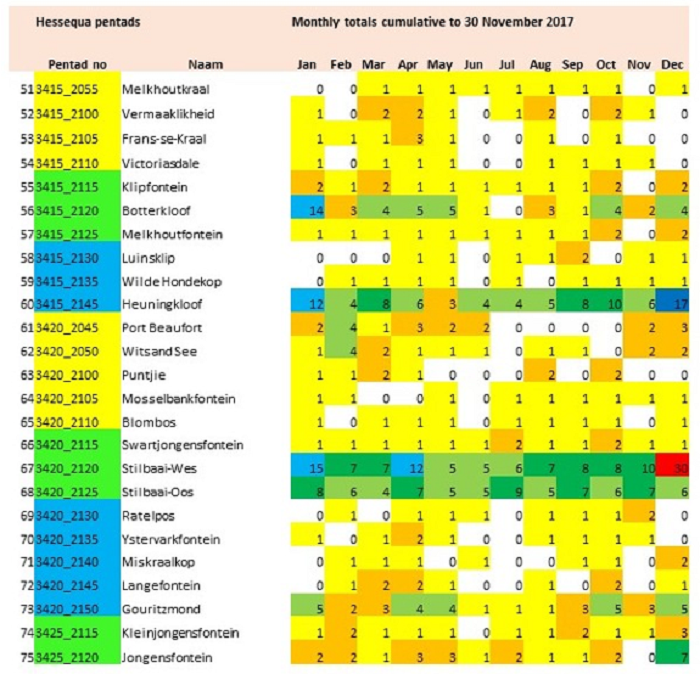
The main focus of the Calvinia BioBash team was the bird atlas. The map below shows the coverage “before” and “after” the BioBash. Do a bit of visual exercise to see the difference between the two maps. You discover that a lot of pentads were atlased for the first time. Changes in colour between the two coverage maps show pentads which received additional checklists.
Our accommodation at ‘n Handvol Gruis was in the pentad with the blue star. Inspection of the “before” and “after” maps shows that a lot of pentads were atlased for the first time, and the changes in colour in the between the two coverage maps show pentads that received second, third or fourth checklists.
We also worked on enriching the Virtual Museum database (http://vmus.adu.org.za). Between us, with Zenobia van Dyk and myself being chief contributors, we added about 500 records to the various sections of the Virtual Museum. We highlight a few of the records.
Roads are essential for the collection of biodiversity data. They made doing the Calvinia BioBash feasible! However, their direct impact on biodiversity is generally negative. Especially snakes and mammals become road casualties. Taking a photo and uploading it to the Virtual Museum means that the wasted animal is not a total dead loss, unless of course it is the last representative of the species in the district. The largely dried out Puff Adder above became a valuable point in the distribution map for this species. Believe it or not, this is the first ever formal record of Puff Adder in the quarter degree grid cell 3119BC, which lies immediately west of Calvinia, and with good roads. So even the published reptile atlas does not have Puff Adder for this grid cell. In fact, this grid cell has only had five records of reptiles, representing four species since 1980. The Puff Adder is the fourth! To see the map of this grid cell, and a list of the four species, go to http://vmus.adu.org.za/vm_locus_map.php?vm=ReptileMAP&locus=3119BC! This illustrates how much fieldwork still needs to be done!
Taking a photo of this young Yellow Canary was greatly facilitated by the fence. It won’t win a prize for elegance, but the fence is a wonderful perch for BirdPix records. It is in the BirdPix section of the Virtual Museum at http://vmus.adu.org.za/?vm=BirdPix-64300.
Overall, fences are not a positive for biodiversity. But they are a big plus for BirdPixers. We are not bothered by the aesthetics of an ugly barbed wire fence. All we need are images in which the bird is easily identified. Fences provide great perches where we can take photos of a whole bird rather than a partly obscured bird.
Poles are substitute trees, enabling crows to breed in places that would otherwise have very few nest sites. This pair of Cape Crows in in BirdPix at http://vmus.adu.org.za/?vm=BirdPix-64432.
Across much of the arid Karoo, trees are rare. Poles, and the wires between them, provide elevated perches for many bird predators, creating hunting opportunities that never existed before. Predators have an unobstructed view of the ground below, a luxury unavailable in pristine conditions, when the best hunting perch might be a shrub. It is likely that a more serious problem with poles is that they provide substitute trees for the nests of crows. Poles have enabled crows to spread into arid areas of South Africa. Controlling crows is not going to make any long-term difference, because there are plenty of spare crows to take the places of any that are culled. Removing poles might have a long-term impact.
The Hantam River has carved itself a gorge, and a trickle of water was still flowing. The gravel road winds down to the bottom, and the meeting of road and stream makes a fabulous spot to go dragon hunting.
Dragonflies are mostly associated with water. But the Hantam area is arid. So one would expect dragonflies to be as rare as rocking horse droppings. But there are isolated patches of water. There is a wonderful “watersplash” where the Hantam River crosses the gravel road in a remarkable gorge a few kilometres north of the farm Kaalplek. This was probably the best spot in quarter degree grid cell 3119BB for dragonflies. To see the map of this grid cell, and a list of the six species of dragonflies and damselflies recorded here, go to http://vmus.adu.org.za/vm_locus_map.php?vm=OdonataMAP&locus=3119BB!
Perhaps the prize record of the Calvinia BioBash for OdonataMAP was this Vagrant Emperor, photographed by Zenobia van Dyk. In the entire Northern Cape, this was the fifth quarter degree grid cell in which the species has been recorded.
Dragonflies of the family called the “emperors” are hard to shoot. They seldom stop for a rest! So we have to do our best to photograph them in flight. This is, without uncertainty, a Vagrant Emperor, because of the blue “saddle” at the back end of the thorax. Zenobia van Dyk shot this dragon at a small spring-fed lake near the farm house ‘n Handvol Gruis. This image is curated at http://vmus.adu.org.za/?vm=OdonataMAP-59371.
This neat brown and white butterfly is a Namaqua Bar. It was photographed in the Hamtam River gorge. It is the first record of this species in this quarter degree grid cell. So it was not shown here in the butterfly atlas! It is curated here: http://vmus.adu.org.za/?vm=LepiMAP-662313.
At the start of the expedition, the number of butterfly species recorded in quarter degree grid cell 3919BB was seven. Four species were photographed and uploaded. Three were the extremely common and almost ubiquitous African Monarch, Painted Lady and Common Meadow White. The fourth, Namaqua Bar (in the photo above) was identified by Fanie Rautenbach, LepiMAP expert panel. Astonishingly, all four species were new to the grid cell! See the list below! You get the up-to-date list by clicking on http://vmus.adu.org.za/vm_locus_map.php?vm=LepiMAP&locus=3119BB, and you can see if any additional records have been added.
This is the list of LepiMAP species provided by the Virtual Museum for grid cell 3119BB.
All the columns are easy, but the last two need some explanation. The column headed “Last recorded” provides the most recent date on which a species was recorded in the Virtual Museum. This provides you with an insight into how urgently each species needs to be “refreshed”. Ideally, you should download this list before you go into the field, and choose a set of priority species for “refreshment”. A species which was last recorded 10 years or longer ago is definitely needing a new record to confirm that it is still present in the grid cell. Even a three-year old record needs refreshing. If you have a series of photos for a grid cell, upload them all. Don’t worry if some the “Last recorded” dates are recent. Any species which are not already on the list are especially valuable and important!
Every entry in the final column reads “Records”, in blue. Click on this and you will discover it is a link to all the records of the species in the grid cell (including any from before 1980!). But it won’t work here, because this is a photograph of the table! It is fascinating to be empowered to see when the records were made, and who the observer was.
The entrance to the farm Kaalplek
Kaalplek has been mentioned a couple of times above. To an English-speaking South African, with a modest grip on Afrikaans, this translates into “the place where you walk around naked”. This very literal translation does not capture the intended meaning: “the place which is barren and treeless.” The Hantam is a tough area to be a farmer, and the droughts of the past few years have resulted in many of the farms being totally abandoned.
Salome Willemse and Zenobia van Dyk did a reconnaissance trip to find the accommodation and test the roads for quality, and were part of the expedition, from 8 to 12 November 2018. Alan Collett and Tino Herselman traveled west from the Karoo to participate. Eric Hermann traveled north from Hopefield, and I came from Cape Town. We are all grateful to Salome for her coordination and leadership of the expedition. The Calvinia BioBash was citizen science at its best.
Loftie-Eaton M, Underhill LG, and Navarro R. 2018. OdonataMAP – Progress report on the Atlas of the Dragonflies and Damselflies of Africa – 2016/17 and 2017/18. Biodiversity Observations 9.13:1-10
OdonataMAP – Progress report on the Atlas of the Dragonflies and Damselflies of Africa – 2016/17 and 2017/18
Megan Loftie-Eaton
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; Biodiversity and Development Institute, 25 Old Farm Road, Rondebosch, 7700 South Africa
Les G Underhill
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; Biodiversity and Development Institute, 25 Old Farm Road, Rondebosch, 7700 South Africa
Rene Navarro
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; FitzPatrick Institute of African Ornithology, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa
Abstract
This paper reports progress with OdonataMAP, the Atlas of Dragonflies and Damselflies of Africa, for the two-year period 1 July 2016 to 30 June 2018. During the two-year review period, the database for the project grew by 30,423 records to 52,257, starting from 22,809 records collected between 2010 and June 2016. Submissions were made from 25 African countries. In six of the nine provinces of South Africa, the number of OdonataMAP records for the province more than doubled. The provinces in which the number of records were not doubled were Gauteng (44% of records made during reporting period), Free State and North West (both 46%). Five observers contributed more than 1000 records over the two-year period, and a further 10 between 500 and 999 records. The total number of observers for the two-year period was 529, compared with 295 in the 2010-16 period. One of the important success of OdonataMAP during the review period was to increase the number of observers, and to reduce the project’s dependence on a small number of citizen scientists.
What is OdonataMAP?
OdonataMAP is the Atlas of Dragonflies and Damselflies of Africa. It was launched in 2010; the first record was uploaded into the OdonataMAP database on 22 September 2010. At the time, the project objectives were (1) “to map the current distribution of the insect Order Odonata, i.e. dragonflies and damselflies, occurring in Africa” and (2) “to serve as a repository of all existing distribution data for this group.”
OdonataMAP provides up-to-date distributions of the dragonflies and damselflies, a critical component of addressing their own conservation priorities, and also the conservation of freshwater ecosystems. The project will provide a valuable input to a revision of the Red List status of each species.
Besides the distribution maps, OdonataMAP aims to make a contribution to understanding the seasonal phenology of these species. Documenting and quantifying changes in seasonality are a critical component of understanding the impacts of climate change on biodiversity (Hassel et al. 2007, Bush et al. 2013).
Thus OdonataMAP aims not only to mainstream the conservation of the Odonata, but also the freshwater habitat on which they (and we) depend. Besides its own value for the conservation of Odonata, this atlas has the potential to influence government policy on the use of water resources, especially for the rural poor. There is no other taxon which has the potential to be so politically prominent in this way.
The report by Underhill et al. (2016) summarized what OdonataMAP had achieved during the period 22 September 2010 until 30 June 2016. The database then contained 22,809 records. The end of June is a natural splitting point for the Odonata year, because it is midwinter in the southern hemisphere and fieldwork is at its lowest ebb. This report focuses on the growth of the database for the two “years”, from 1 July 2016 to 30 June 2017, and from 1July 2017 to 30 June 2018.
The Odonata Database of Africa (ODA) is an open access database developed by a JRS-funded project (Clausnitzer et al. 2012, Dijkstra 2016). This database contains 121,121 records of the distribution of dragonflies and damselflies across Africa and it includes most of the museum specimen records for the region.This database became available online during the last months of 2016, where it is known as African Dragonflies and Damselflies Online (ADDO) (http://addo.adu.org.za/). ADDO is a collaboration between the Department of Conservation Ecology and Entomology (University of Stellenbosch) and the ADU (University of Cape Town). Although the two databases are separate, search queries made to the OdonataMAP database can include a search of the Odonata Database of Africa. This collaboration, completed during the two-year reporting period, represents a major consolidation of data resources.
How many records were submitted to OdonataMAP in the period July 2016 to June 2018, and where in Africa did they come from?
For the years July 2016 to June 2017 and from June 2017 to July 2018, OdonataMAP gained 11,091 and 18,357 records respectively, a total of 30,423 new records, bring the grand total since the start of the project in 2010 to 52,257 records (Table 1). This is an increase from 22,809 records in June 2016, thus 56.4% of the OdonataMAP database has been contributed in the two years under review (Table 1).
Table 1: Annual totals (1 July to 30 June of following calendar year) of submissions of dragonflies and dragonflies to OdonataMAP
Year (July to June)
Number of submissions
Cumulative totals
Cumulative percentage
2010/11
349
349
0.7
2011/12
951
1300
2.5
2012/13
4000
5300
10.1
2013/14
5074
10374
19.9
2014/15
3997
14371
27.5
2015/16
8438
22809
43.6
2016/17
11091
33900
64.9
2017/18
18357
52257
100.0
The records submitted during the two year reporting period came from 25 African countries; the total number of countries for which records have been submitted is now 32 (Table 2). Outside of South Africa, most of the records for the reporting period came from Namibia (491) and more than 100 records from 11 other African countries (Namibia, Botswana, Zambia, Nigeria, Malawi, Angola, Kenya, Mozambique, Zimbabwe, Swaziland, and Sudan). Two records were submitted from countries in the Middle East (Table 2).
Table 2: Numbers of submissions of dragonflies and damselflies to OdonataMAP from countries of Africa (and the Middle East) prior to and during the two-year reporting period
Country
2010-2016
2016/17
2017/18
Total
Angola
14
164
125
303
Benin
5
NA
NA
5
Botswana
213
137
262
612
Democratic Republic of Congo
4
63
44
111
Iraq
NA
NA
1
1
Israel
NA
NA
1
1
Egypt
3
NA
NA
3
Ethiopia
90
NA
NA
90
Gambia
4
NA
NA
4
Ghana
13
68
NA
81
Kenya
82
220
55
357
Lesotho
5
8
NA
13
Liberia
8
8
NA
16
Madagascar
44
NA
NA
44
Malawi
441
144
201
786
Mauritius
1
NA
NA
1
Mozambique
157
19
192
368
Namibia
145
132
359
636
Nigeria
75
320
35
430
Republic of the Congo
7
12
NA
19
Reunion
NA
NA
7
7
Rwanda
6
19
NA
25
Senegal
7
14
NA
21
Seychelles
6
NA
2
8
Sierra Leone
35
76
NA
111
Somalia
1
NA
NA
1
South Africa
20339
9475
17347
47161
Sudan
13
NA
118
131
Swaziland
556
60
89
705
Tanzania
51
3
13
67
Togo
3
NA
NA
3
Uganda
73
29
52
154
Zambia
259
26
334
619
Zimbabwe
144
77
94
315
Total
22804
11074
19331
53209
In six of the nine provinces of South Africa, more than 50% of the total number of records had been submitted in the two-year reporting period (Table 3). The largest percentage increase was for the Western Cape, for which 6,635 records were submitted, 74% of the of the total number of OdonataMAP records for the province. The provinces in which the number of records were not doubled were Gauteng (44% of records during reporting period), Free State and North West (both 46%) (Table 3). Overall, for South Africa as a whole, 57% of records were submitted during the reporting period (Table 3).
Table 3: Numbers of submissions of dragonflies and damselflies to OdonataMAP from the nine provinces of South Africa prior to and during the two-year reporting period. The percentage of records for each province during the reporting period is provided in the final column
Province
2010-2016
2016/17
2017/18
Total
Percentage of records 2016-2018
Eastern Cape
1454
591
1730
3775
61.5
Free State
607
280
241
1128
46.2
Gauteng
1257
529
460
2246
44.0
KwaZulu-Natal
7597
3162
4755
15514
51.0
Limpopo
2845
988
2378
6211
54.2
Mpumalanga
2777
744
2601
6122
54.6
North West
788
266
413
1467
46.3
Northern Cape
566
429
412
1407
59.8
Western Cape
2275
2433
4202
8910
74.5
Total
20166
9422
17192
46780
56.9
What were the outcomes of the Shoot the Dragons Weeks of the past two summers?
OdonataMAP hosted a series of 10 “Shoot the Dragons Weeks” in the summers of the reporting period, three in 2016/17 and seven in 2017/18 (Table 4). They ran from the Saturday of one week to the Sunday of the following week, so that they included two weekends. Their purpose was to promote participation in OdonataMAP, and to maintain momentum in data collection and submission. There is a full description of the results of the very first week (Underhill et al. 2016a). The total number of records submitted during the Shoot the Dragons Weeks was 9,270, so that 30% of the records during the reporting period were submitted during the 10 Shoot the Dragons Weeks. To put this into context, the 90 days of the 10 Weeks represents about 20% of the period of the year when the Odonata are most active. This provides a coarse measure of their effectiveness. Shoot the Dragons Weeks will be repeated in the 2018/19 summer.
Table 4: Shoot the Dragons Weeks for the summers of 2016/17 and 2017/18
Week
Start date
End date
Records
Observers
Countries
Taxa
2016/17
2016/17
1
26 November 2016
4 December 2016
1200
61
8
116
2
21 January 2017
29 January 2017
1384
58
10
120
3
1 April 2017
9 April 2017
634
50
5
83
2017/18
2017/18
1
7 October 2017
15 October 2017
590
48
12
91
2
28 October 2017
5 November 2017
884
54
6
95
3
25 November 2017
3 December 2017
701
53
10
90
4
13 January 2018
21 January 2018
1234
66
9
117
5
10 February 2018
18 February 2018
1073
53
8
112
6
10 March 2018
18 March 2018
745
42
6
92
7
7 April 2018
15 April 2018
825
60
5
92
How is the number of OdonataMAP observers growing?
Five observers contributed more than 1000 records over the two-year period, and a further 10 between 500 and 999 records (Table 5). The most prolific observer contributed 6% to the total number of records; by contrast, in the 2010-16 report (Underhill et al. 2016b), the top two observers contributed 19% and 15% of the total number of records. The total number of observers for the two-year period was 529, compared with 295 in the 2010-16 period (Underhill et al. 2016b). One of the important success of OdonataMAP during the review period was to increase the number of observers, and to reduce the project’s dependence on a small number of citizen scientists.
Table 5: OdonataMAP observers who submitted more than 150 records for the reporting period (1 July 2016 to 30 June 2018)
Observer
Records
Ryan M Tippett
2082
Jean Hirons
1683
Christopher Peter Small
1663
Corrie du Toit
1505
Richard Alan Johnstone
1233
Andries Petrus de Vries & Joey de Vries
895
Desire Darling & Gregg Darling
829
Alan Manson
797
Maritza Van Rensburg
742
Andre Marais
623
Sharon Stanton & Heleen Louw
609
Altha Liebenberg
606
Christopher Willis
575
Alf Taylor & Hilary Harrison
568
Alicia Culverwell
547
Niall Perrins
495
John H Wilkinson
493
Rob Dickinson
468
Bensch Gert & Juan-Pierre Antunes
458
Ilse Hulme
429
Dawie Kleynhans & Sarieta Kleynhans
415
Diana Russell
365
Juan-Pierre Antunes & Gert Bensch
313
Christopher JH Hines
290
Gary Brown
288
Phillip Nieuwoudt
264
Jacobus (Lappies) Labuschagne
262
Pieter La Grange
260
Wilna Steenkamp
255
Bensch Gert
247
Bernardine Alice Altenroxel
227
David Kennedy
209
Juan-Pierre Antunes
207
Zenobia van Dyk
205
Riëtte Griesel
197
Norman Barrett
193
Sharon Basel
192
Katharina Reddig
188
Herb Kageler
181
Sharon Stanton
176
Laban Njoroge
171
Waterberg Team 2017 (Andries Petrus de Vries & Joey de Vries)
167
Pieter Cronje
165
Growing numbers of records generated increased workloads for the expert panel. This is a group of volunteers who either undertake identifications from scratch or confirm the identifications made by the observers. Over the reporting period, the load has been shared mainly between John Wilkinson, Ryan Tippett, Sharon Stanton, Alan Manson, Bertie Brink and Lappies Labushagne. Warwick Tarboton remains the anchor to whom difficult records get referred, and the expert panel has also consulted K-D Dijkstra from time to time.
What are a few of the most remarkable records submitted to OdonataMAP during the past two years?
Figure 1: OdonataMAP record 50330 submitted to OdonataMAP by Christopher Hines from Angola. This is, in all likelihood, a new species. (http://vmus.adu.org.za/?vm=OdonataMAP50330)
Commenting on the original posting of the photo in the Facebook group called Dragonflies and Damselflies of Southern Africa, Jens Kipping, authority on the Odonata of Angola, wrote: “Holy moly, Christopher Hines! First, I thought that somebody from the South American or Asian group sent a picture accidentally. I cannot believe that this is from Angola. I do not have any clue what this libellulid is! It looks a bit like Rhyothemis but also, from the body, a bit like a Palpopleura. This might even be a new genus.” KD Dijkstra, taxonomic authority on African Odonata, subsequently analysed the images, and considered it likely that this might prove to be a new species in the genus Trithemis.
An unexpected and dramatic range expansion occurred during the reporting period. The Ceres Streamjack (also known as Spesbona) Spesbona angusta was described in 1863, but was thought for several decades to be extinct, having not been recorded since 1920. The streams in the area near Ceres, Western Cape, where it had been observed in 1920, had been radically transformed and many no longer flowed due to over-extraction of water for the fruit industry. It was rediscovered in November 2003, when a population was found along the Dutoitsrivier, which flows into the Theewaterskloof Dam, near Villiersdorp, Western Cape, South Africa. This locality is 60 km distant from the original Ceres locality. There are multiple sightings in this immediate Theewaterskloof Dam area (OdonataMAP database), and the IUCN-defined Area of Occupancy is 24 km2 (Samways 2018). For more than a decade, this was thought that this was the only locality where the species occurred. Then, on 10 October 2017, citizen scientist Jean Hirons caused a massive surprise when she photographed the species at a locality near Sedgefield, 330 km due east of the Theewaterskloof site (Figure 2). This raises the obvious question: does it occur at a series of intermediate localities, in suitable habitats along the mountain ranges that link these two isolated sites? The likely answer is yes, because an inspection of Figure 3 of Underhill et al. (2018) reveals that, while fieldwork in the areas of the two known localities of the Ceres Streamjack have been reasonably intensive, the intervening area has been poorly covered by fieldwork, and is regarded as a priority area for future expeditions.
Figure 2: This record, by Jean Hirons, of a Ceres Streamjack (Spesbona) Spesbona angusta in Sedgefield, Western Cape, was 330 km east of the only known locality for the species. (http: //vmus.adu.org.za/?vm=OdonataMAP-35883)
What are the take-home messages?
In a nutshell, the OdonataMAP project grew rapidly in the two-year period under review. From a public-interest perspective, there can be no doubt that the Odonata have been transformed from being the taxon of focus for a minuscule group of enthusiasts, to becoming quite substantial. It is likely that the size of the dragonfly/damselfly community is in the process of overtaking that of the butterfly community, if it has not done so already. It is still far smaller than the bird community, but that is challenge that OdonataMAP is taking on.
Multiple factors have played a role in this growth. Emerging at roughly the same time, they have interacted with each other, and reinforced each other: (1) the publication of the superb fieldguide (Tarboton & Tarboton 2015); (2) the excellently managed Dragonflies and Damselflies of Southern Africa group on Facebook; and (3) the sturdy Virtual Museum platform for uploading images into a long-term database; (4) the award of funding to the Animal Demography Unit at UCT by the JRS Biodiversity Foundation, Seattle, USA.
Growing the broad civil society interest in the Odonata is part of the strategy for meeting the next challenge. This is in fact the challenge set by the JRS Biodiversity Foundation: “How do we get the data into use? How do we mainstream the dragonflies and damselflies so that the OdonataMAP data become serious components of conservation policy making, of environmental impact of assessments, and the thinking of politicians and civil servants?” Our report on the Odonata of the Kruger National Park was an experimental step in that direction (Underhill et al. 2018).
Acknowledgements
John Wilkinson, Alan Manson and Lappies Labuschagne and others made helpful suggestions. We acknowledge funding from the JRS Biodiversity Foundation, Seattle, USA. But above all, we celebrate the amazing contributions made by two teams of citizen scientists: the fieldworkers who contribute the records and the expert panel who undertake the identifications.
References
Clausnitzer V, Dijkstra K-DB, Koch R, Boudot J-P, Darwall WRT, Kipping J, Samraoui B, Samways MJ, Simaika JP, Suhling F 2012. Focus on African freshwaters: hotspots of dragonfly diversity and conservation concern. Frontiers in Ecology and the Environment 10: 129-134
Dijkstra, K-DB 2016. African Dragonflies and Damselflies Online. (Version 1 July 2016). Available online at http://addo.adu.org.za .
Samways MJ, Tarboton W 2006. Rediscovery of Metacnemis angusta (Selys, 1863) in the Western Cape, South Africa (Zygoptera: Matycnemididae). Odonatologica 35: 375-378
Tarboton W, Tarboton M 2015. A Guide to Dragonflies and Damselflies of South Africa. Struik Nature, Cape Town.
Underhill LG, Manson AD, Labuschagne JP, Tippett JM 2016a. Shoot the Dragons Week, Round 1: OdonataMAP grows by 1,200 records. Biodiversity Observations 7.100: 1-14. Available online at https://journals.uct.ac.za/index.php/BO/article/view/393/434
Underhill LG, Navarro R, Manson AD, Labuschagne JP, Tarboton WR 2016b. OdonataMAP: progress report on the atlas of the dragonflies and damselflies of Africa, 2010-2016. Biodiversity Observations 7.47: 1-10. Available online at https://journals.uct.ac.za/index.php/BO/article/view/340
Underhill LG, Loftie-Eaton M, Navarro R 2018. Odonata of the Kruger National Park. Biodiversity Observations 9.11:1-16.
de Swardt DH. 2018. Notes on the seasonal occurrence of Sickle-winged Chats Cercomela sinuata in the central Free State with notes on their seasonality. Biodiversity Observations 9.12:1-5
Notes on the seasonal occurrence of Sickle-winged Chats Cercomela sinuata in the central Free State with notes on their seasonality
Dawid H de Swardt
Department of Ornithology, National Museum, P O Box 266, Bloemfontein 9300, South Africa
Dean (2005) described the status of the Sickle-winged Chat Cercomela sinuata as resident with some altitudinal movements from the Drakensberg escarpment to the lower Lesotho and KwaZulu-Natal areas. These represents mostly C. s. hypernephela (Dean 2005) and this species’ geographic variations are not discussed in Chittenden et al. (2012). Brown & Barnes (1984) first described these movements on the Alpine belt on the Drakensberg escarpment to adjacent lower altitudes. Harrison (1997) described their status (during SABAP1 surveys) with lower summer reporting rates and with winter increases in the central western parts of their distribution (Zone 3 on SABAP1 distribution map) which includes most of the central and western Free State. Harrison (1997) also suggested a seasonal movement into and out of the winter rainfall succulent Karoo areas.
During fieldwork for both the SABAP1 and current SABAP2 bird atlas projects (and BIRP projects), I observed winter seasonal influxes of Sickle-winged Chats in the central parts of the Free State. These winter increases were mostly observed during the April – August period when this species was observed in abundance in its preferred habitat. It is still uncertain from where these individuals originate, but the SABAP2 data will possibly support / explain the suggestion of seasonal influx into and out of the winter rainfall areas of the succulent Karoo to the central parts of their range (Harrison 1997).
The aim of this paper is to analyse the winter / summer seasonal distribution of the Sickle-winged Chat and to analyse its monthly occurrences in certain selected degree grid cells (DGCs). This paper will focus mainly on the central and western Free State and areas in the Western Cape and southern Eastern Cape where seasonality is also suspected to occur.
The distribution of Sickle-winged Chats is presented as winter (April – September) (Figure 1a) and summer (October – March) (Figure 1b) patterns and the higher reporting rates during winter in the central Free State are clearly noticeable (see explanations in Underhill & Brooks 2016). Also the map shows some differences in seasonal reporting rates in the Western Cape and southern Eastern Cape areas.
Figure 1: SABAP2 distribution maps for the winter (a) and summer (b) months for the Sickle-winged Chat, downloaded on 19 April 2017. The higher winter reporting rates (a) in the central parts of Free Sate differ noticeably from the lower reporting rates of the summer map (b). The detailed interpretation of this map is provided by Underhill & Brooks (2016) and see text. There are no data from pentads shaded turquoise. Pentads with white dots have fewer than four checklists but the species was not recorded. Pentads with four or more checklists are either shaded white, species not recorded, or in colour, with shades based on reporting rate: yellow 0-7.9%, orange 7.9-17.0%, light green 17.0-28.9%, dark green 28.9-44.8%, light blue 44.8-64.9% and dark blue 64.9-100%.
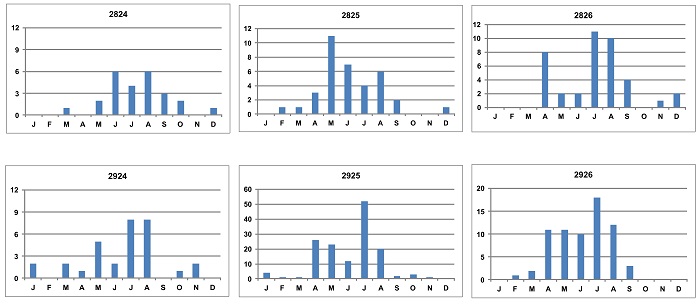
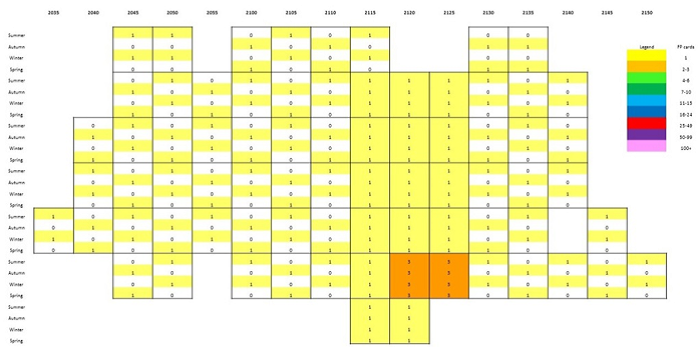
In the central and western parts of the Free State peak reporting rates were recorded in 6 DGCs mostly from April / May to August / September (Figure 2). Lower summer occurrences were also noted. In the more southern Free State (areas along the Gariep River) in DGCs 3024, 3025 and 3026 (not shown in Figure 2), Sickle-winged Chats were recorded in all months, with some summer peaks. During the Birds in Reserves Project (BIRP) which started in 1992 (after SABAP1), there was also a March / April – September peak in reporting rates recorded at Soetdoring and Willem Pretorius Nature Reserves (De Swardt 2000), which supports the SABAP2 data presented in Figure 1a & b. Also, BIRP data from Tussen-die-Riviere Game Reserve near Bethulie (which is along the Gariep River) show a summer peak from August – November.
Figure 2: Bar graphs of degree grid cells (DGCs) in the central and western Free State showing the monthly seasonal patterns of occurrence of Sickle-winged Chats. Note the absence or low reporting rates during the summer months (October – March).
In parts of the Western Cape (see Figure 1a & b) winter to early summer peaks were recorded during SABAP2 (Figure 3). In DGCs 3024, 3025 and 3026 low to high winter peaks were complimented by higher summer reporting rates to early summer reporting peaks during SABAP2 (not shown in Figure 3). In DGC 3324 (not shown in Figures 2 or 3), in the southern Eastern Cape, Sickle-winged Chats were recorded during all months of the year with an August peak, but the seasonal difference in Figure 1a & b is also noticeable. More distribution data are still needed in several DGCs in the Sickle-winged Chat distributional range, especially in low coverage areas.
Figure 3: Bar graphs of degree grid cells (DGCs) in the Western Cape areas showing the monthly seasonal patterns of occurrence of Sickle-winged Chats. Note the higher reporting rates during the summer months and also late winter peaks in some areas.
This is the first time that the seasonal occurrence of Sickle-winged Chats has been analysed based on SABAP2 distributional data, and supports the suggestion that this species is not only resident in certain areas of its range, but that seasonal movements do occur (see Dean 2005, Harrison 1997). It is also the first attempt to analyse the winter influx of Sickle-winged Chats to the central and western Free State based on SABAP2 data, personal observations during the various atlas projects, and earlier published observations (see De Swardt 2000). This study also supports Harrison’s (1997) speculation that movement of this species does occur into and out of the winter rainfall succulent Karoo and the Western Cape areas. Oatley (2017) further recorded Sickle-winged Chats as breeding in the higher altitudes of Lesotho and recorded the species on rare occasions in the Drakensberg foothills in winter. Oatley (2017) further noted and suggested that this species’ migration is mostly westwards towards the Free State, which is supported by the SABAP2 data and personal observations in this study.
A total of 36 Sickle-winged Chats have been ringed so far, most of them in Lesotho, KwaZulu-Natal (Sani Pass areas) and in the Eastern Cape. There is only one ringing record from the Glen area in the central Free State. Also, only one recapture has been obtained with no movement recorded for the individual. A colour ringing study on this species is needed to unravel their seasonal occurrence in the central Free State and other parts of their range.
Acknowledgements
Thanks due to all the Citizen Scientists who submitted SABAP2 cards with Sickle-winged Chats on their lists, and to Les Underhill for producing the Sickle-winged Chat summer / winter maps for this manuscript.
References
Brown CJ, Barnes PR 1984. Birds of the Natal Alpine belt. Lammergeyer 33: 1-13.
Chittenden H, Allan DG, Weiersbye I 2012. Roberts geographic variation of southern African birds. John Voelcker Bird Book Fund: Cape Town.
De Swardt DH 2000. The birds of Soetdoring Nature Reserve and adjacent areas, central Free State. BirdLife South Africa guide pp. 1-40. BirdLife South Africa: Johannesburg.
Dean WRJ 2005. Sickle-winged Chat Cercomelia sinuata In: Hockey PAR, Dean WRJ, Ryan PG. (Eds). Roberts – Birds of Southern Africa (VIIth Ed) Cape Town: The Trustees of the John Voelcker Bird Book Fund. pp. 952-953.
Harrison JA 1997. Sickle-winged Chat. In: Harrison, J.A., Allan, D.G, Underhill, L.G., Herremans, M., Tree, A.J., Parker, V. & Brown, C.J. (Eds.). The atlas of southern African birds. Vol. 2, pp. 180-181. Johannesburg: BirdLife South Africa.
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; Biodiversity and Development Institute, 25 Old Farm Road, Rondebosch, 7700 South Africa
Megan Loftie-Eaton
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; Biodiversity and Development Institute, 25 Old Farm Road, Rondebosch, 7700 South Africa
Rene Navarro
Animal Demography Unit, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa; FitzPatrick Institute of African Ornithology, Department of Biological Sciences, University of Cape Town, Rondebosch, 7701 South Africa
Abstract
The number of species of dragonflies and damselflies recorded in the Kruger National, South Africa, was 103 in April 2018. This figure was based on a database containing 2,817 records of Odonata, made since 1980, from the 52 quarter degree grid cells which intersect with the Kruger National Park. Records were available for 41 of the 52 grid cells. The most frequently recorded species were Red-veined Dropwing Trithemis arteriosa (167 records) and the Orange-veined Dropwing Trithemis kirbyi (144 records), both recorded in 33 grid cells, and Southern Banded Groundling Brachythemis leucosticta (175 records) and Broad Scarlet (141 records) both in 29 grid cells. Based on records up to April 2018, the median date of the most recent record for species was September 2017, so that half of the 103 species had been recorded during summer 2017/18. This report could be used to motivate the proclamation of the river and wetland systems of the Kruger National Park as a ‘Wetland of International Importance’ in terms of the Ramsar Convention. Two-thirds of the Odonata of South Africa, and one-eighth of the Odonata of Africa, have been recorded in the Kruger National Park.
Dragonflies and damselflies are important indicators of water quality and ecosystem health (Figure 1)
Figure 1. A Barbet Percher Diplacodes luminans recorded by Craig Peter on the southern bank of the Luvuvhu River near Pafuri, Kruger National Park. OdonataMAP record 46300 http://vmus.adu.org.za/?vm=OdonataMAP-46300
Introduction
This document is experimental. It aims to provide a model for the presentation of biodiversity data that can be used by managers and policy makers, by researchers, and by citizen scientists. For these groups of people it aims (1) to provide a snapshot, at a point in time, of the quality and volume of data available for a locality, and (2) aims to provide links to the relevant databases, so they have access to useful summaries of the ongoing data collection effort. In this case the locality is the Kruger National Park, South Africa, and the component of biodiversity under consideration is the Odonata, the dragonflies and the damselflies.
We are looking for suggestions that will improve the usefulness of this product. We are primarily hoping that this review will be on value to managers and policy makers, so it is their information needs which we primarily want to meet.
Study area: Kruger National Park, South Africa
The Kruger National Park (KNP) is the flagship national park of South Africa. The KNP, located in the north-eastern corner of South Africa, was established as a government reserve in 1898 and became South Africa’s first national park in 1926. It was first established to control over-hunting and to protect the dwindling number of herbivores in the Lowveld (Stevenson-Hamilton 1993). The KNP is currently nearly two million hectares in size (19,485 km2). It is a national conservation icon of South Africa and considered to be a safe haven for many fauna and flora. KNP is home to approximately 2,000 species of plant, 53 fish, 34 amphibians, 118 reptiles, 517 birds and 147 mammal species (SANParks 2016).
KNP is about 360 km long from north to south, and about 65 km wide on average, from west to east. Its widest point is 90 km (Paynter & Nussey 1986). The park is bordered by the Limpopo River in the north and the Crocodile River in the south, forming natural park boundaries. Several other rivers run through the park from west to east, including the Sabie, Olifants, Letaba and Luvuvhu Rivers. The Lebombo Mountains lie on the eastern park boundary with Mozambique and to the west the KNP is fringed with many other private nature reserves (forming part of the Greater Kruger National Park) and local communities, villages and towns. The park’s altitude ranges from 200 m to 800 m. The highest point is Khandzalive Hill in the south-west of the park near the Berg-en-Dal rest camp (Paynter & Nussey 1986).
The Lowveld, and consequently the KNP, has a subtropical climate. Subtropical climates are characterised by warm, humid summers and mild, dry winters. Summer temperatures can rise above 38 °C. The rainy season starts around November and lasts until May. The driest period is September and October (SANParks 2016).
Data resources
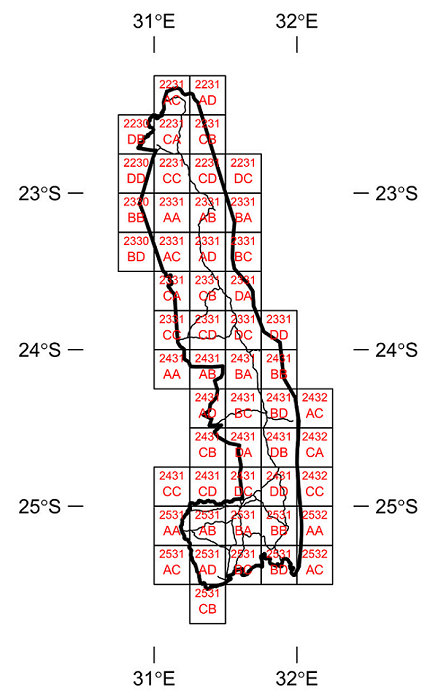
This document provides information related to the Odonata for the Kruger National Park as well as a selection of species distribution maps. It makes use of the open access database developed by a project funded by the JRS Biodiversity Foundation which generated the Odonata Database of Africa (Clausnitzer et al. 2012, Dijkstra 2016, available online as African Dragonflies and Damselflies Online at http://addo.adu.org.za) and the citizen science database generated by the OdonataMAP project (Underhill et al. 2016, available online at http://vmus.adu.org.za). Both databases are open access. This report is based on species recorded in 52 quarter degree grid cells which fall entirely or partly within the KNP (Table 1, Figure 2). Search queries made to the OdonataMAP database can be extended to include a search of the Odonata Database of Africa, which includes almost all of the museum specimen records for the region. This has been done for this report.
Table 1. The codes for the Quarter Degree Grid Cells (QDGC) which fall fully or partly within the Kruger National Park (KNP). The column headed ‘% in KNP’ provides an estimate of the percentage of the QDGC which lies inside the KNP.
QDGC and name
% in KNP
2230DB HAMAKUYA
17
2230DD KA-XIKUNDU
16
2231AC MABILIGWE
44
2231AD PAFURI
12
2231CA PUNDA MARIA
99
2231CB MACHAYIPAN
50
2231CC DZUNDWINI
100
2231CD SHINGOMENI
83
2231DC
0
2330BB SHANGONI
28
2330BD NSAMA
2
2331AA SHIGOMANE
100
2331AB SHINGWIDZI
100
2331AC NALATSI
91
2331AD DZOMBO
100
2331BA SHINGWIDZI (EAST)
16
2331BC KOSTINI
22
2331CA MAHLANGENI
50
2331CB NGODZI
100
2331CC PHALABORWA
35
2331CD MASORINI
100
2331DA SHILOWA
67
2331DC LETABA
97
2331DD GORGE
19
2431AA GRIETJIE
5
2431AB ROODEKRANS
53
2431AD ORPEN
37
2431BA BALULE
100
2431BB BANGU
62
2431BC MASALA
100
2431BD SATARA
96
2431CB MANYELETI
7
2431CC BOSBOKRAND
0
2431CD NEWINGTON
9
2431DA RIPAPE
77
2431DB LINDANDA
100
2431DC SKUKUZA
63
2431DD TSHOKWANA
100
2432AC
1
2432CA
1
2432CC TSHOKWANA
5
2531AA KIEPERSOL
14
2531AB PRETORIUSKOP
100
2531AC WITRIVIER
0
2531AD GUTSHWA
85
2531BA DUBE
100
2531BB ONDER-SABIE
100
2531BC HECTORSPRUIT
67
2531BD KOMATIPOORT
46
2531CB KAAPMUIDEN
3
2532AA ONDER-SABIE
12
2532AC KOMATIPOORT
4
Figure 2. Locations of the 52 Quarter Degree Grid Cells (QDGCs) which intersect with the Kruger National Park. These are listed in Table 1. The naming convention follows the tradition that has been used in South Africa for almost a century. There are 16 QDGCs in a one-degree grid cell. Each one-degree cell is numbered by the coordinates, latitude first, then longitude, of the northwest corner of the cell. The subdivisions of the one-degree grid cell have an alphabetic notation, as shown.
OdonataMAP data for the Kruger National Park
On 23 April 2018, there were 2,817 records of Odonata in the combined database of OdonataMAP and the Odonata Data Base of Africa, recorded since 1980. Of these, 2663 had been identified to species level, and the remainder to genus level. The number of species recorded for the Kruger National Park was 103 species from eight families (Table 2). Within Table 2, the ordering is first alphabetically by family, and then by genus and species.
Table 2. Species of Odonata recorded in the 52 quarter degree grid cells (Table 1) which intersect with the Kruger National Park, South Africa. The cut-off date is 1980; i.e. records prior to this are not included in this analysis. The number of quarter degree grid cells in which each species has been recorded is given, and n refers to the number of records in joint ADDO-OdonataMAP database for the species. The table lists 103 species.
Species code
Family
Scientific name
Common name
Grid cells
n
Most recent record
664070
Aeshnidae
Anaciaeschna triangulifera
Evening Hawker
1
1
2017/09/01
664120
Aeshnidae
Anax ephippiger
Vagrant Emperor
6
7
2014/03/29
664140
Aeshnidae
Anax imperator
Blue Emperor
20
41
2016/10/06
664170
Aeshnidae
Anax speratus
(Eastern) Orange Emperor
6
9
2012/12/06
664180
Aeshnidae
Anax tristis
Black Emperor
4
5
2012/12/06
664320
Aeshnidae
Gynacantha manderica
Little Duskhawker
1
1
2002/01/01
664470
Aeshnidae
Pinheyschna subpupillata
Stream Hawker
1
1
2001/01/01
660580
Calopterygidae
Phaon iridipennis
Glistening Demoiselle
14
34
2018/01/28
661180
Chlorocyphidae
Platycypha caligata
Dancing Jewel
10
28
2018/01/27
662330
Coenagrionidae
Africallagma glaucum
Swamp Bluet
5
8
2013/02/18
662460
Coenagrionidae
Agriocnemis exilis
Little Wisp
1
1
2001/01/01
662470
Coenagrionidae
Agriocnemis falcifera
White-masked Wisp
1
1
2013/02/18
662630
Coenagrionidae
Azuragrion nigridorsum
Sailing Bluet
11
22
2018/02/03
662720
Coenagrionidae
Ceriagrion glabrum
Common Citril
21
68
2018/03/16
662790
Coenagrionidae
Ceriagrion suave
Suave Citril
1
2
2014/03/29
663100
Coenagrionidae
Ischnura senegalensis
Tropical Bluetail
18
56
2018/03/16
663670
Coenagrionidae
Pseudagrion acaciae
Acacia Sprite
18
52
2017/08/07
663710
Coenagrionidae
Pseudagrion coeleste
Catshead Sprite
2
2
2015/10/13
663720
Coenagrionidae
Pseudagrion commoniae
Black Sprite
18
43
2014/05/04
663360
Coenagrionidae
Pseudagrion gamblesi
Great Sprite
7
15
2016/07/12
663410
Coenagrionidae
Pseudagrion hageni
Painted Sprite
4
10
2017/01/27
663780
Coenagrionidae
Pseudagrion hamoni
Swarthy Sprite
23
91
2018/03/16
663460
Coenagrionidae
Pseudagrion kersteni
Powder-faced Sprite
16
35
2017/01/23
663820
Coenagrionidae
Pseudagrion massaicum
Masai Sprite
17
72
2017/12/01
663560
Coenagrionidae
Pseudagrion salisburyense
Slate Sprite
7
10
2015/11/23
663870
Coenagrionidae
Pseudagrion sjoestedti
Variable Sprite
6
11
2014/03/28
663880
Coenagrionidae
Pseudagrion sublacteum
Cherry-eye Sprite
19
58
2018/01/27
663890
Coenagrionidae
Pseudagrion sudanicum
Blue-sided Sprite
5
17
2017/10/07
664550
Gomphidae
Cerato-gomphus pictus
Common Thorntail
1
1
2006/12/12
664640
Gomphidae
Creni-gomphus hartmanni
Clubbed Talontail
9
16
2016/05/17
664770
Gomphidae
Gomphidia quarrei
Southern Fingertail
3
6
2016/12/27
664830
Gomphidae
Ictino-gomphus ferox
Common Tigertail
16
51
2018/01/28
664880
Gomphidae
Lestino-gomphus angustus
Spined Fairytail
4
5
2012/11/24
665300
Gomphidae
Neuro-gomphus zambeziensis
Zambezi Siphontail
4
10
2017/12/08
665480
Gomphidae
Notogomphus praetorius
Yellowjack Longleg
1
1
2013/01/25
665640
Gomphidae
Onycho-gomphus supinus
Lined Claspertail
1
1
2002/01/01
665740
Gomphidae
Paragomphus cognatus
Rock Hooktail
4
6
2017/01/23
665780
Gomphidae
Paragomphus elpidius
Corkscrew Hooktail
10
15
2018/01/27
665790
Gomphidae
Paragomphus genei
Common Hooktail
17
36
2017/11/18
665840
Gomphidae
Paragomphus magnus
Great Hooktail
5
6
2017/03/20
665890
Gomphidae
Paragomphus sabicus
Flapper Hooktail
5
9
2018/01/14
666070
Gomphidae
Phyllogom-phus selysi
Bold Leaftail
3
4
2018/02/20
660410
Lestidae
Lestes pallidus
Pallid Spreadwing
7
8
2017/11/21
660360
Lestidae
Lestes plagiatus
Highland Spreadwing
5
10
2018/01/30
660330
Lestidae
Lestes tridens
Spotted Spreadwing
2
2
2002/01/01
660370
Lestidae
Lestes uncifer
Sickle Spreadwing
5
5
2018/03/23
660300
Lestidae
Lestes virgatus
Smoky Spreadwing
1
2
2013/02/18
666750
Libellulidae
Acisoma inflatum
Stout Pintail
3
3
2017/01/20
666770
Libellulidae
Acisoma variegatum
Slender Pintail
4
12
2018/03/16
666920
Libellulidae
Aethriamanta rezia
Pygmy Basker
1
1
1992/01/01
667020
Libellulidae
Brachythemis lacustris
Red Groundling
16
50
2018/01/27
667030
Libellulidae
Brachythemis leucosticta
Southern Banded Groundling
29
175
2018/03/22
667060
Libellulidae
Bradinopyga cornuta
Horned Rockdweller
10
15
2018/03/16
667090
Libellulidae
Chalcostephia flavifrons
Inspector
2
32
2018/02/04
667130
Libellulidae
Crocothemis erythraea
Broad Scarlet
29
143
2018/03/16
667140
Libellulidae
Crocothemis sanguinolenta
Little Scarlet
7
12
2015/10/09
667200
Libellulidae
Diplacodes lefebvrii
Black Percher
15
42
2018/03/22
667210
Libellulidae
Diplacodes luminans
Barbet Percher
14
22
2018/03/22
667380
Libellulidae
Hemistigma albipunctum
African Piedspot
5
11
2017/09/01
667690
Libellulidae
Nesciothemis farinosa
Eastern Blacktail
21
105
2018/02/04
667730
Libellulidae
Notiothemis jonesi
Eastern Forest-watcher
3
4
2012/12/06
667760
Libellulidae
Olpogastra lugubris
Bottletail
4
5
2017/11/29
667780
Libellulidae
Orthetrum abbotti
Little Skimmer
2
2
2012/12/06
667830
Libellulidae
Orthetrum brachiale
Banded Skimmer
1
1
2014/03/29
667860
Libellulidae
Orthetrum caffrum
Two-striped Skimmer
1
1
2007/01/27
667900
Libellulidae
Orthetrum chrysostigma
Epaulet Skimmer
24
91
2018/01/28
667930
Libellulidae
Orthetrum hintzi
Dark-shouldered Skimmer
3
4
2017/11/01
667940
Libellulidae
Orthetrum icteromelas
Spectacled Skimmer
1
1
2011/03/07
667950
Libellulidae
Orthetrum julia
Julia Skimmer
13
45
2018/02/04
668000
Libellulidae
Orthetrum machadoi
Highland Skimmer
3
27
2018/02/20
668110
Libellulidae
Orthetrum stemmale
Bold Skimmer
9
36
2018/02/20
668120
Libellulidae
Orthetrum trinacria
Long Skimmer
19
43
2018/01/30
668180
Libellulidae
Palpopleura deceptor
Deceptive Widow
4
16
2014/05/04
668190
Libellulidae
Palpopleura jucunda
Yellow-veined Widow
5
7
2014/05/02
668200
Libellulidae
Palpopleura lucia
Lucia Widow
22
68
2018/02/20
668210
Libellulidae
Palpopleura portia
Portia Widow
10
21
2017/11/26
668230
Libellulidae
Pantala flavescens
Wandering Glider
22
61
2018/01/28
668370
Libellulidae
Rhyothemis semihyalina
Phantom Flutterer
8
15
2016/04/11
668420
Libellulidae
Sympetrum fonscolombii
Red-veined Darter or Nomad
4
4
2014/04/27
668540
Libellulidae
Tetrathemis polleni
Black-splashed Elf
8
15
2018/02/03
668600
Libellulidae
Tholymis tillarga
Twister
1
1
2012/02/10
668620
Libellulidae
Tramea basilaris
Keyhole Glider
13
17
2018/01/31
668630
Libellulidae
Tramea limbata
Ferruginous Glider
10
13
2014/11/18
668740
Libellulidae
Trithemis aconita
Halfshade Dropwing
6
10
2018/01/30
668660
Libellulidae
Trithemis annulata
Violet Dropwing
16
114
2018/03/23
668670
Libellulidae
Trithemis arteriosa
Red-veined Dropwing
33
167
2018/01/30
668800
Libellulidae
Trithemis donaldsoni
Denim Dropwing
7
8
2017/01/23
668870
Libellulidae
Trithemis dorsalis
Highland Dropwing
2
2
2016/02/02
668890
Libellulidae
Trithemis furva
Navy Dropwing
3
6
2017/01/27
669120
Libellulidae
Trithemis kirbyi
Orange-winged Dropwing
33
144
2018/03/17
668900
Libellulidae
Trithemis pluvialis
Russet Dropwing
3
9
2016/06/07
669080
Libellulidae
Trithemis stictica
Jaunty Dropwing
5
5
2015/03/14
669130
Libellulidae
Trithemis werneri
Elegant Dropwing
7
26
2018/03/15
669180
Libellulidae
Urothemis assignata
Red Basker
10
22
2016/05/18
669190
Libellulidae
Urothemis edwardsii
Blue Basker
8
13
2016/04/11
669250
Libellulidae
Zygonoides fuelleborni
Southern Riverking
9
13
2016/12/27
669390
Libellulidae
Zygonyx natalensis
Blue Cascader
7
12
2017/11/29
669420
Libellulidae
Zygonyx torridus
Ringed Cascader
16
24
2014/05/04
666420
Macromiidae
Phyllomacro-mia contumax
Two-banded Cruiser
7
9
2014/05/01
666620
Macromiidae
Phyllomacro-mia picta
Darting Cruiser
8
10
2017/12/01
661480
Platycnemididae
Allocnemis leucosticta
Goldtail
2
3
2015/03/14
661810
Platycnemididae
Elattoneura glauca
Common Threadtail
13
37
2018/01/30
661640
Platycnemididae
Mesocnemis singularis
Common (Forest/ Savanna) Riverjack
9
13
2017/01/23
The Red-veined Dropwing Trithemis arteriosa (167 records) and the Orange-veined Dropwing Trithemis kirbyi (144 records) were both recorded in 33 of the quarter degree grid cells of the Kruger National Park, and Southern Banded Groundling Brachythemis leucosticta (175 records) and Broad Scarlet (141 records) in 29 grid cells (Table 2). These four dragonflies were the most widely distributed species.
The dataset is commendably “young” (final column of Table 2). Based on records up to April 2018, the median date of the most recent record for species was September 2017. In other words, half of the 103 species had been recorded during the most recent eight-month period, i.e. in summer 2017/18. The lower quartile was in March 2015, indicating that three-quarters of the species have been recorded in the most recent three years.
Special attention needs to be focused on “refreshing” the records of the species in oldest quartile; in this case, it is species not recorded since 2015. Of species recorded since 1980, nine have not been recorded for more than 10 years, i.e. prior to 2008 (Table 2). Common Thorntail Ceratogomphus pictus was last recorded on 12 December 2006, Two-striped Skimmer Orthetrum caffrum was last recorded on 27 January 2007, and Spectacled Skimmer Orthetrum icteromelas was last recorded on 7 March 2011 (Table 2). For a further six species, only the year of the last record is available: Pygmy Basker Aethriamanta rezia (1992), Steam Hawker Pinheyschna subpupillata and Little Wisp Agriocnemis exilis (2001), and Spotted Spreadwing Lestes tridens, Lined Claspertail Onychogomphus supinus and Little Duskhawker Gynacantha maderica (2002). Eight of these nine species have been recorded only once in the Kruger National Park since 1980, and the Spotted Spreadwing twice (Table 2). The presence of these species needs careful evaluation. Six species were last recorded in 2012, four in 2013, and 11 in 2014.
Of the species in the “oldest” quartile, three had been recorded in more than 10 grid cells: Black Sprite Pseudagrion commoniae (18 grid cells, 43 records), Ringed Cascader Zygonyx torridus (16 grid cells, 24 records) and Ferruginous Glider Tramea limbata (10 grid cells, 13 records) (Table 2). The current status of these three species should be investigated. They were last recorded in 2014 (Table 2).
The maximum number of species of Odonata in any of the 52 quarter degree grid cells of the Kruger National Park was 60 (Figure 3). The median was 12 species. Eleven grid cells had no records of Odonata. Most of these grid cells have only a small percentage of their area within the Kruger National Park, and those on the eastern edge of the park are mostly in Mozambique and are virtually inaccessible even from within that country (Peter Lawson pers. comm.). If these grid cells are excluded from the calculation, the median number of species per grid cell is 21.
Figure 3. Species richness of the Odonata (dragonflies and damselflies) in the Quarter Degree Grid Cells (QDGC) which intersect with the Kruger National Park. Refer to Figure 2 and Table 1 for the naming conventions of each QDGC.
Table 3. The number of species recorded in each of the 41 quarter degree grid cells intersecting with the Kruger National Park which have Odonata data. The quarter degree grid cell code is provided, the number of species recorded, and the number of records of these species. Because of the difficulty of making identifications of species with certainty from photographs, some records are identified to genus or family. The final column gives the total number of records for the grid cell in the database.
QDGC
No. of species
Records identified to species level
Number of taxa
Total number of records
2230DB HAMAKUYA
36
58
37
59
2231AC MABILIGWE
42
69
46
75
2231AD PAFURI
48
122
48
122
2231CA PUNDA MARIA
30
77
31
78
2231CC DZUNDWINI
20
46
21
48
2231CD SHINGOMENI
15
20
16
21
2331AB SHINGWIDZI
28
77
30
79
2331AD DZOMBO
3
7
4
8
2331BA SHINGWIDZI (OOS)
9
11
12
15
2331BC KOSTINI
11
18
12
19
2331CA MAHLANGENI
8
8
8
8
2331CB NGODZI
25
112
27
117
2331CC PHALABORWA
8
16
9
17
2331CD MASORINI
7
10
8
12
2331DA SHILOWA
4
5
5
6
2331DC LETABA
45
194
48
197
2331DD GORGE
4
9
4
9
2431AA GRIETJIE
42
258
48
291
2431AB ROODEKRANS
3
3
3
3
2431AD ORPEN
3
3
5
5
2431BA BALULE
29
88
33
95
2431BB BANGU
21
27
22
29
2431BC MASALA
13
16
13
16
2431BD SATARA
25
52
27
54
2431CB MANYELETI
20
35
24
40
2431CC BOSBOKRAND
9
10
10
11
2431CD NEWINGTON
47
154
53
167
2431DA RIPAPE
5
6
5
6
2431DB LINDANDA
4
4
4
4
2431DC SKUKUZA
60
244
66
253
2431DD TSHOKWANA
55
133
58
138
2531AA KIEPERSOL
59
211
64
218
2531AB PRETORIUSKOP
8
13
9
14
2531AC WITRIVIER
25
49
28
54
2531AD GUTSHWA
21
37
24
47
2531BA DUBE
12
16
12
16
2531BB ONDER-SABIE
44
101
48
105
2531BC HECTORSPRUIT
54
152
58
158
2531BD KOMATIPOORT
50
370
57
394
2531CB KAAPMUIDEN
51
114
51
114
2532AA ONDER-SABIE
1
1
1
1
What is immediately clear from Figure 3 is that the species richness within the park appears to be spatially uneven, with no strong geographical pattern (such as a decrease from north to south, or east to west). Figure 3 is the result of two confounded processes: the fieldwork behaviour of the observers (technically, the observer process) and the truth on the ground (the biological process). Although there is likely to be variation in the number of species per grid cell, due to the uneven distribution of suitable wetland habitat for Odonata, the true variation is unlikely to be as large as depicted in Figure 3. The challenge for fieldwork in the Kruger National Park is that the dragonflies and damselflies share their habitat with Nile Crocodiles Crocodylus niloticus, Common Hippopotamuses Hippopotamus amphibious and other life-threatening animals. As a result, and in accordance with the SANParks code of conduct for visitors to the Kruger National Park, most river systems and wetlands are out of bounds; this makes consistent recording of especially the damselflies difficult, and introduces a bias into the data.
In spite of this, it is probably feasible, now that the unevenness of the observer effort is displayed (Figure 3), that the citizen scientists who are the primary contributors of data to OdonataMAP will find a way to reduce as much of the bias introduced by the observer process as feasible.
Distribution maps, generated in May 2018, are provided for a sample of four species, selected to illustrate various levels of occurrence in the Kruger National Park (Figures 4 and 5). Up-to-date distribution maps (i.e. for use in the future) for all species can be obtained from the following link:
Figure 4. Distribution maps for the Red-veined Dropwing *Trithemis arteriosa* and the Epaulet Skimmer *Orthretrum chrysostigma* in the Kruger National Park and surrounding areas. The Red-veined Dropwing has been recorded in 33 of the 52 QDGCs which intersect with the Kruger National Park, the most widespread species, and the Epaulet Skimmer in 24. The distribution beyond the Kruger National Park is displayed on these maps.
This gives the map for the species with species code number 668670, the Red-veined Dropwing (Figure 4). The species codes are provided in the first column of Table 2.
Figure 5. Distribution maps for the Violet Dropwing *Trithemis annulata* and the Darting Cruiser *Phyllomacromia picta* in the Kruger National Park and surrounding areas. The Violet Dropwing has been recorded in 16 of the 52 QDGCs which intersect with the Kruger National Park and the Darting Cruiser in eight. The distribution beyond the Kruger National Park is displayed in these maps.
Up-to-date lists of the species recorded in a quarter degree grid cell can be obtained from the following link. The list of grid cell codes is provided in Table 1. In the link below, replace the “locus” with the code for the QDGC required (consult also Figure 2):
These lists are constructed to the same format as that of Table 2, including the provision of the most recent record of each species in the grid cell. The list can be copied and pasted to Excel, where they can be sorted and manipulated as needed. These list include numbers of records for which the expert panel was unable to make an identification to “species” level.
Earlier studies of the Odonata of the Kruger National Park
Three key papers have been written which focus on the Odonata of the Kruger National Park (Table 4). The first was a list of 21 species recorded by Balinsky (1965). Three decades late, Clark & Samways (1994) generated a list of 80 species, based on three sources: the 21 species by Balinsky (1965), their own list of 59 species from along the Sabie River, and a list of 61 species generated by “other collectors” in the period between Balinsky’s fieldwork and their own. Clark & Samways (1994) noted that there were three species which had only been recorded by Balinsky (1965); these are included in the list of Table 2; i.e. their presence has been confirmed. They also noted that there were 11 species which were only on the list made by “other collectors”. All except one of these species is included in Table 2; the exception is Cryptic Syphontail Neurogomphus vicinus, a species which has only been recorded from its type locality in the Democratic Republic of Congo (Schouteden 1934); so this species represents an error.
Table 4. Papers which focus on reviews of the Odonata (dragonflies and damselflies) of the Kruger National Park.
Reference
Summary
Balinsky BI 1965. A preliminary list of the dragonflies (Odonata) of the Kruger National Park. Koedoe 8: 95-96
Based on a total of 84 specimens collected in the Kruger National Park, this paper provided a list of 21 species. At the end of this Balinsky provides reasons why ‘the present list includes only a small fraction of the dragonfly species occurring in the Park.’
Clark TE, Samways MJ 1994. An inventory of the damselflies and dragonflies (Odonata) of the Kruger National Park, with three new South African records. African Entomology 2: 61-64
Using the list by Balinsky (1065) as baseline, made three decades previously, records made by other researchers, and their own data from a study of the Odonata along the Sabie River, the authors extended the list to 81.
Clark TE, Samways MJ 1996. Dragonflies (Odonata) as indicators of biotope quality in the Kruger National Park, South Africa. Journal of Applied Ecology 33: 1001-1012
This paper demonstrated how the Odonata can be used to undertake biomonitoring of the Sabie River. Ten ‘biotopes’ (waterbody types) were identified, and the Odonata species characteristic of each were identified using multivariate statistical methods.
It is remarkable how the list of Odonata species has grown in the past five decades, from 21 to 80 and currently 103 (Tables 2 and 4). It is even more remarkable that two-thirds of South Africa’s species of Odonata (Tarboton & Tarboton 2015) have been recorded in the quarter degree grid cells which intersect with the Kruger National Park.
Conclusions and recommendations
This paper has aimed to highlight the contents of the OdonataMAP database, in relation to the Kruger National Park, providing a snapshot from April-May 2018. But it also provides the tools to enable users (1) to obtain up-to-date species distribution maps for the dragonflies and damselflies of the Kruger National Park, and (2) to obtain up-to-date species lists on the scale of the quarter degree grid cell. These maps and lists are extracted “on the fly” from the live database for the project when the queries are made.
These distribution maps and species lists can only be comprehensive if the OdonataMAP database contains the entire knowledge base. One of the concerns of the second decade of the 21st century is the proliferation of a variety of biodiversity data collection initiatives. This serves to split (and confuse) the citizen scientist community, which is in any event small, and to generate a diversity of databases which do not communicate with each other, and ultimately result in products such as those produced within this system being incomplete. The OdonataMAP database (supplemented by the Odonata Data Base of Africa (Clausnitzer et al. 2012, Dijkstra 2016) which contains the overwhelming majority of specimen records in museum collections, and the observations of taxon specialists), is currently the most reliable and up to date database of the Odonata in Africa, and growing rapidly (Underhill et al. 2016).
We are seeking suggestions for additional resources which would be perceived valuable. For example, both managers and citizen scientists might be interested in a species map which shows the time elapsed, in appropriate units, since the species was recorded in each grid cell. From a management perspective, if a pattern emerges, it is a warning that a species is becoming hard to locate in an area, and that it might be going extinct there. From the citizen scientist perspective, this knowledge provides an incentive of which species need to be “refreshed” in each grid cell. Another example might be a table which shows the median date of the records from each quarter degree grid cell. From both management and citizen scientist perspective, this provides guidance as to where observer effort should be focused.
How can these data be used for annual monitoring of Odonata in the Kruger National Park? Because the bulk of the records are made by citizen scientists it is difficult to impose a strict protocol on data collection. However, with some ingenuity, it ought to be feasible to encourage citizen scientists, cumulatively, to visit as many grid cells as they are able, and to use these data some form of occupancy modelling to estimate changes in distribution and seasonality through time. There is a natural annual pattern to the occurrence of adult dragonflies and damselflies, with a winter lull. This annual cycle can be used to plan citizen scientist data collection strategies for the upcoming summer.
Given the hazards of doing fieldwork in the presence of dangerous wild animals (and in fact the understandable prohibition on this by SANParks for citizen scientists), it is not going to be feasible to undertake a complete survey of the distribution of dragonflies and damselflies of the Kruger National Park. It is therefore sensible to think in terms of using species distribution models to achieve this (Elith & Leathwick 2009, Franklin 2009, Guisan et al. 2013). Although in this paper, the data have been summarized in terms of quarter degree grid cells, the overwhelming majority of the individual records are georeferenced. This means that it is possible to use a species distribution model system such as MaxEnt to generate plausible distributions of species (Elith et al. 2011).
Finally, this paper ends up effectively being a motivation for the proclamation of the Kruger National Park, or at least its river and wetland systems, as a “Wetland of International Importance” in terms of the Ramsar Convention (Ramsar Convention on Wetlands 2016). About two-thirds of the Odonata ever recorded in South Africa have been recorded here. About one-eighth of the Odonata of the continent of Africa have been recorded in the Kruger National Park.
Acknowledgements
The primary acknowledgement needs to be attributed to the people who collected the data, both professional entomologists and citizen scientists. We acknowledge funding support from the JRS Biodiversity Foundation. Many people commented on earlier drafts.
References
Balinsky BI 1965. A preliminary list of the dragonflies (Odonata) of the Kruger National Park. Koedoe 8: 95-96.
Clark TE, Samways MJ 1994. An inventory of the damselflies and dragonflies (Odonata) of the Kruger National Park, with three new South African records. African Entomology 2: 61-64.
Clark TE, Samways MJ 1996. Dragonflies (Odonata) as indicators of biotope quality in the Kruger National Park, South Africa. Journal of Applied Ecology 33: 1001-1012.
Clausnitzer V, Dijkstra K-DB, Koch R, Boudot J-P, Darwall WRT, Kipping J, Samraoui B, Samways MJ, Simaika JP, Suhling F 2012. Focus on African freshwaters: hotspots of dragonfly diversity and conservation concern. Frontiers in Ecology and the Environment 10: 129-134
Dijkstra, K-DB 2016. African Dragonflies and Damselflies Online. (Version 1 July 2016). Available online at http://addo.adu.org.za .
Elith J, Leathwick JR 2009. Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution and Systematics 40: 677-697.
Elith J, Phillips SJ, Hastie T, Dud KM, Chee YE, Yates CJ 2011. A statistical explanation of MaxEnt for ecologists. Diversity and Distributions 17: 43-57.
Franklin J 2009. Mapping Species Distributions: Spatial Inference and Prediction. Cambridge University Press, Cambridge, UK.
Guisan A, Tingley R, Baumgartner JB, Naujokaitis-Lewis I, Sutcliffe PR, Tulloch AIT, Regan TJ, Brotons L, McDonald-Madden E, Mantyka-Pringle C, Martin TG, Rhodes JR, Maggini R, Setterfield SA, Elith J, Schwartz MW, Wintle BA, Broennimann O, Austin M, Ferrier S, Kearney MR, Possingham HP, Buckley, YM 2013. Predicting species distributions for conservation decisions. Ecological Letters 16:1424-1435.
Paynter D, Nussey W 1986. Die Krugerwildtuin in Woord en Beeld. Macmillan Suid-Afrika, Johannesburg.
Ramsar Convention on Wetlands 2016. An Introduction to the Ramsar Convention on Wetlands. Ramsar Convention Secretariat, Gland, Switzerland.
SANParks 2016. South African National Parks. Kruger National Park: Introduction. Retrieved from https://www.sanparks.org/parks/kruger/ on 5 October 2016.
Schouteden H 1934. Les Odonata des Congo Belge. Annales Musée Congo Belge. Zoologie Serie 3 Section 2. Catalogue Raisonnés de la Fauna Entomolgique du Congo Belge Tome 3, Fascicule 1: 1-84.
Stevenson-Hamilton J 1993. South African Eden: The Kruger National Park 1902-1946. Struik Publishers, Cape Town.
Tarboton W, Tarboton M 2015. A Guide to Dragonflies and Damselflies of South Africa. Struik Nature, Cape Town.
Underhill LG, Navarro R, Manson AD, Labuschagne JP, Tarboton WR 2016. OdonataMAP: progress report on the atlas of the dragonflies and damselflies of Africa, 2010-2016. Biodiversity Observations 7.47: 1-10. Available online at http://bo.adu.org.za/content.php?id=240
Elith J, Leathwick JR 2009a. Species distribution models: ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution and Systematics 40: 677-697.
Franklin J 2009. Mapping Species Distributions: Spatial Inference and Prediction. Cambridge University Press, Cambridge, UK.
How frequently can the species in the photograph be identified?
We asked this question “What’s the value of a photo?” of seven of the taxa in the Virtual Museum (http://vmus.adu.org.za). We looked at scorpions, lacewings, birds, frogs, reptiles, butterflies, and dragonflies and damselflies.
What happens is this. Citizen scientists upload their photographs into the Virtual Museum. A record can consist of one to three photographs. The second and third photographs are usually used to show different angles on the animal, or they can be used to show habitat. They will know that the record is butterfly, and that it ought to be submitted to LepiMAP, and that the scorpions go to ScorpionMAP, etc. But they don’t need to know what species they are uploading (although they can submit their identification, or even tentative identification). For each of these groups of species, there is an expert panel consisting of specialists, often citizen scientists themselves. These experts do their best to make an identification to species (or even subspecies level for some butterflies and reptiles). But they are careful, and if the identification cannot be made with certainty to species level, they allocate a genus, and sometimes even a family. This mostly happens when the photo is a bit blurred, or if the critical ID feature is not shown. And some species can only be identified when the species is captured, and the critical diagnostic features, often the genitals, are examined with a magnifying glass! The identifications are displayed for all to see and examine on the Virtual Museum website; as a result, the members of the expert panels tend to err on the side of caution.
Let’s work our way through the seven groups. For each group of species, we will count the number of records submitted, and calculate the percentage which have been identified to species level. Each group is illustrated with a recently uploaded record from the Virtual Museum.
This is Opistophthalmus gigas. At this stage the scorpions only have scientific names. There is a great need to make them more accessible to everyone by allocating English common names! The photograph was taken by Luke Kemp in Namibia. The ScorpionMAP details are available at http://vmus.adu.org.za/?vm=ScorpionMAP-3490
Scorpions. We expected the scorpions to be tricky. But of the first 3,259 scorpions to be submitted to ScorpionMAP, 3,259 have been identified to species. That is an incredible 98.6%.
This is one of the “green lacewings”. It is identified to family Chrysopidae. To see more detail, go to LacewingMAP and look at http://vmus.adu.org.za/?vm=LacewingMAP-15501. This photograph was uploaded from Swaziland by Kate Braun. Mervyn Mansell comments: “Unfortunately it is not possible to identify any closer. It is quite difficult to distinguish between the various green lacewing species without a close examination of the wing venation and the male genitalia. The females of the different species are difficult to separate even if wing venation is clear. Some species are, however, characteristic and can be distinguished from photos, This is is possibly one of the members of the genus Chrysoperla”
Lacewings. We knew the lacewings would be really really difficult. For the “green lacewings”, the Chrysopidae, many species need to be dissected to identify them! In spite of the problems, Mervyn Mansell, the lacewing specialist, has identified 1,556 of the 2,380 records uploaded to LacewingMAP to species. That is a commendable 65.4%. Even for the green lacewings, the ID-to-species rate is 35%. That is far higher than Mervyn anticipated when LacewingMAP started.
The beautiful Common Moorhen was photographed today, 2 November 2018, and the photo immediately uploaded by Adriette Benade from near Alexander Bay in the Northern Cape, South Africa. The BirdPix record is permanently curated at http://vmus.adu.org.za/?vm=BirdPix-63485
Birds. 61,657 bird records have been submitted to BirdPix. 61,518 have been identified to species. That is 99.8%. That is the highest rate of all. BirdPix has the potential to make a huge contribution to the African Bird Atlas. Wherever you are in Africa, take photos of birds and upload them. The chances of the species being identified are overwhelming!
Frogs. The frogs are potentially difficult to identify from photographs. Sometimes the key ID features can only be seen if there is a series of photos from different angles. The number of records submitted is 8,221, of which 7,898 have been identified to species. That is 96.1%. Given the difficulties, that is remarkably high.
Reptiles. The number of sets of reptile photos uploaded to ReptileMAP is 19,980. Many species of reptile are cryptic, and identification involves counting scales, etc. So it is not self-evident that the rate of identification from photos will be large. But the success rate in the Virtual Museum is an astonishing 98.8%. 19,736 of the records have been have identified to species or subspecies level.
Ryan Tippett uploaded this exquisite Eastern Sorrel Copper. He took the photo in the Clocolan District of the Free State. It was the fourth record of this species from this grid cell. That sounds pretty boring. Until you grasp that it “refreshed” the most recent previous record, made in December 1995. That’s 23 years ago. Refreshing old records is incredibly important, because it provides evidence that a species still occurs at a site. Details of this record are at http://vmus.adu.org.za/?vm=LepiMAP-661549
Butterflies. The butterflies constitute the largest component of the Virtual Museum. There are 88,892 sets of up to three photographs. 95.3% have been identified to species or sub-species level. There are six families of butterflies, and four are unproblematic! 88% of the butterflies identified to family or genus level come from two families: the Lycaenidae (the blues and the coppers) and the Hesperiidae (the skippers). These are the two tricky families with lots of lookalike species.
Citizen Scientist Alan Manson has made a massive contribution to OdonataMAP, especially from KwaZulu-Natal. Once upon a time, he was a birder, but then he saw the light, and got hooked on the dragonflies and damselflies. He took this photograph of a Violet Dropwing near Komatipoort in Mpumalanga. Full details are at http:// http://vmus.adu.org.za/?vm=OdonataMAP-58739
Dragonflies and Damselflies (the Odonata). The OdonataMAP database is growing fast. It now consists of 56,039 records. This is another tricky group, in which identification depends on subtle features. In spite of this, the expert panel has identified 51,291 records to species level. That’s an amazing 91.5%.
Take home message. 95.6% success rate in identifying to species!
The take home message. The question we asked at the start was “How frequently can the species in the photograph be identified?” Across the seven groups of species considered here, by November 2018, a total of 240,428 records have been submitted to the Virtual Museum. That is almost a quarter of a million records. Of these, 229,927 have been identified by one of the expert panels to species (or subspecies) level. That is an impressive 95.6%. For six of the seven groups, the identification-to-species rate exceeds 90%. The lacewings get dragged down by the “green lacewings”. The take home message has to be that the value of a photo record to biodiversity mapping at the species level, as done by the Virtual Museum, is massive.
What motivates this blog on industrial biodiversity? Well, the BDI took occupation of its unit in Epping Industria I on Monday this past week. Director Pete Laver is doing some spectacular innovations. Watch this space.
Meanwhile we are exploring our new neighbourhood. It’s a mix of industrial warehouses and factories. Lots of hard surfaces: roads, roofs and paving. But the sheer number of birds is impressive.
The most common species is House Sparrow. This is the male of a pair with a nest behind the fire alarm. What happens when the alarm goes off?House Sparrows everywhere. This rust inspector is on the job.Most razor wire never actually gets to serve its function of keeping intruders out. Here is an alternative use for razor wire. Providing this House Sparrow with a perch in the early morning sun. Although Factory Sparrows (sorry, House Sparrows) are abundant, they do not manage to totally exclude Cape Sparrows.Rock Doves (feral pigeons) are also almost ubiquitous. This concentration must be finding something special on the stones! You need to remove the speck of dirt from your beautiful sheen.A comfortable spot for a preen.The Rock Doves share this space with three other members of the pigeon-dove family. This industrial Laughing Dove is closing its eyes while the southeaster ruffles its feathers.The Red-eyed Dove has found a daytime use for street-lighting. There IS a little bit of green in this picture. This Speckled Pigeon was finding food on the tar.The third most abundant species is, like the House Sparrow and Rock Dove, an feral alien. The only green in this picture is the sheen on wing of the Common Starling. There’s an indigenous starling too. This Red-winged Starling has found a little snack.
Even in this environment there are people who care for their biodiversity. People, who in their own way, are “connected to nature.” Here are three containers of water. They have rocks in them so that the birds can perch and drink (and so that the southeaster doesn’t blow them away).Epping Industria I, established in the 1940s, is what is known as a novel ecosystem. A space so utterly transformed that it cannot serve any conservation function. Is that really true? If the biodiversity can be used to “connect to nature” the thousands of people who work here, then it can serve a purpose. We live in the “Anthropocene”, the period in which human activity is the dominant influence on the environment and on biodiversity. Ultimately, the only way we are going to achieve biodiversity conservation is by ranking up the concept of “connectedness to nature.” And BDI will try, in its way, to achieve this in Epping Industria I.Here are two species in Epping which don’t have to compete with alien, feral “cousins.” This Cape Wagtail is hiding in the shadow of a security fence ….. and this, with its grey collar, is a Cape Canary … By the time we get to write Industrial Biodiversity 2, the BDI logo will be in the frame above the door of Unit 4. (** but we have moved from here **)
All the photos of birds are preserved for posterity in the BirdPix section of the Virtual Museum.
Industrial Biodiversity 2 deals with the Black River as it flows under the N1 and into Paarden Island.
Dragons do exist, and they’ve been around for over 300 million years! They might not breathe fire, but they do have six legs, four wings, and extremely keen eyesight. These mini dragons are carnivorous insects known as dragonflies (Anisoptera) and damselflies (Zygoptera), belonging to the insect Order Odonata. OdonataMAP, the Atlas of African Odonata, funded by the JRS Biodiversity Foundation is a project in the Animal Demography Unit’s Virtual Museum aiming to: (1) map the current distribution of the dragonflies and damselflies occurring in Africa; and (2) to serve as a repository of all existing distribution data for this group in the geographic extent of the project so that we can better understand their conservation priorities.
The first Shoot the Dragons Week for the 2018 Spring/Summer season kicked off on 22 September 2018. OdonataMAPpers were out and about, armed with their cameras, smiles and enthusiasm, to see how many dragonflies and damselflies they could snap and map. These beautiful insects are important monitors of water quality. They are sensitive to environmental change and play key roles in both terrestrial and aquatic habitats. They are predators as both nymphs and adults, feeding on a variety of prey including nuisance species such as mosquitoes and biting flies. Spending most of their lives underwater in rivers, streams, ponds, and lakes, their presence in aquatic environments is an excellent measure of water quality as they require clean water to thrive.
For the Week (which ran from 22 to 30 September 2018) a total 724 dragons were snapped, mapped and uploaded to the OdonataMAP database at http://vmus.adu.org.za. Records came in from Botswana, Kenya, Namibia, South Africa, Zambia and Zimbabwe! For South Africa, most of the records came from KwaZulu-Natal Province (173).
Diana Russell mapped the most dragons for the Shoot the Dragons Week with 80 records, followed by Norman Barrett (76) and Richard Johnstone (41). This is amazing! A massive thank you also goes to each and everyone that contributed records during the week. Every single record counts. Of the 724 records that were submitted (which covers 73 different taxa), 78,5% have been identified already, thanks to the super OdonataMAP expert panel.
The dragon that was snapped and mapped most commonly during the Week was the Red-veined Darter Sympetrum fonscolombii, with 81 records, followed by the Tropical Bluetail Ischnura senegalensis (52) and Black Percher Diplacodes lefebvrii (34). There were 15 species for which more than 10 records were submitted during the Shoot the Dragons Week.
Bernardine Altenroxel has recently graduated cum laude with a BSc(Hons) in Geography from the University of South Africa (UNISA). So, with her studies behind her for the time being, BDI sent her a new assignment, with a series of questions relating to citizen science. Bernie has contributed more than 4,000 records to the Virtual Museum; insects are her main interest with 3,000 contributions to LepiMAP and 500 to OdonataMAP.
Degrees from UNISA are underpinned by great feats of determination and persistence. Congratulations to citizen scientist Bernadine Altenroxel on the award of a BSc(Hons) in May this year
Bernardine Altenroxel does “disaster management”, the business of turning chaos into order. She says: “If there is anything my line of work has taught me, it is to never take a single day for granted. Any given day has the potential to descend into utter chaos – from accidents, to fires, to floods. If disaster management has taught me about bringing order back from disorder, biomapping has taught me about the need for balance and harmony in the natural environment. One speaks to the other, for without balance and harmony in the natural environment, disorder will inevitably follow; within disorder, balance and harmony need to be restored.”
BDI: Bernie, how did you become a citizen scientist? What was the catalyst that got you going?
In 2009, I had been going through a really rough time and felt the need to get away on my own for a few hours to clear my head. Lekgalameetse Nature Reserve was, and remains, one of my favourite places and this seemed like the ideal place for me to gain clarity on what I should do with my life. There had been profound and abrupt changes in my life which I needed to grasp, and the tranquillity of Lekgalameetse was sure to help me think clearly … plus provide me with the opportunity to test out my recently bought digital SLR camera. On a bright mid-winters day in 2009, I entered Lekgalameetse and to my absolute delight found the solitude I so craved. Besides the Reserve staff, there was not another person that I encountered whilst there. Little did I know it then, but this day would serve as a catalyst which would see me embark on exciting journey of learning and discovery, and entry into the exciting world of citizen science.
The Malta Waterfall at Lekgalameetse Nature Reserve where my journey into the world of citizen science originated.
As I sat beside the Malta Waterfall, deep in thought, a butterfly lazily drifting around in a shaft of sunlight caught my attention. It was unlike any butterfly I could recall having seen before, and I couldn’t help but watch as it landed on a nearby leaf. Barely breathing, I carefully stood up and tried to take a photograph, but the butterfly lifted off the leaf and started to circle again. Dejected at the missed opportunity, I sat down again, but could not help watching as the butterfly fluttered around and then settled on the end of the same leaf again. Eventually I managed to take a photograph of the butterfly. Little did I know that this would be the first of many happy hours spent photographing butterflies. For many months I puzzled over the identity of the butterfly. I just couldn’t get it off my mind. Internet searches yielded nothing and back then I had no books to use as a reference. This single record would eventually serve as the catalyst which would see me drawn into the world of citizen science. The curiosity over the butterfly would just not go away. Eventually, I submitted the record to a citizen science project and finally found the answer I had been looking for. The butterfly was a common mother-of-pearl butterfly. More butterfly records soon followed.
One of the decisions I had taken back in 2009, as I sat beside the Malta Waterfall, was to study further. Having enrolled with UNISA in 2011 for an environmental management degree, I centred my Ecotourism module project (undertaken in 2014) around the butterflies of Lekgalameetse. While researching the project and searching for data on the butterflies of Lekgalameetse, I came into contact with Megan Loftie-Eaton and Professor Les Underhill. They helped me with the information I required, and invited me to become a contributor towards the Animal Demography Unit’s Virtual Museum. It was under their guidance and constant encouragement that I would see myself fully immersed in the world of citizen science, coming into contact with other people who shared my enthusiasm. Pretty soon, I was noticing more than just the butterflies and started to submit records for more of the Virtual Museum projects. This chance encounter with Megan and Les has led me on a path of discovery which has not only shaped my private life and academic progression (I now hold a Batchelor’s degree in Environmental Management and a BSc Honours degree in Geography), but has also contributed significantly towards my work within disaster management. My whole life has been turned around since 2009. Citizen science has literally helped me to heal and grow.
The butterfly which first drew me into citizen science. It is a mother-of-pearl (Protogoniomorpha parhassus) one of the common butterflies of the region.
BDI: What has been the highlight for you?
The absolute highlight was, quite accidentally, re-discovering the Zimbabwe yellow-banded sapphire butterfly (Iolaus nasisii) in 2015, and subsequently being able to locate and successfully rear the Iolaus nasisii caterpillar. I also stumbled upon a thriving community of cryptic spreadwings (Lestes dissimulans) in an area where they have not previously been observed. There can be no better feeling than being able to contribute towards the scientific understanding of species’ distribution and ultimate conservation.
The newly emerged Zimbabwe yellow-banded sapphire butterfly (Iolaus nasisii). While carrying a bundle of hay for my horse on 12 November 2015, I found a small, colourful butterfly. The butterfly was unfortunately dead, probably having succumbed to one of the heatwaves to have gripped Limpopo during October and November. Being a keen LepiMAPper, and instantly recognising the butterfly as an important find, I scooped it up with my free hand, fed my horse and headed back indoors. (http://vmus.adu.org.za/?vm=LepiMAP-582581)
BDI: How has being a citizen scientist changed your view of the world?
Besides bringing me through a difficult period in my life, citizen science has provided me with a deeper understanding of how different components within the environment come together to make an ecosystem whole and functional, and how easily human activities can totally mess this up. This is something I continuously consider in my work within disaster management.
BDI: What are you still hoping to achieve?
Looking ahead, I am hoping for the opportunity to explore more areas and further contribute towards our understanding of the distribution of different species and how this is changing, especially in the face of climate change. My camera is nearly always part of my baggage. I am often required to travel around for work, and will take my camera with me if I think there may be an opportunity to obtain a few more records. This has delivered some surprises, like finding a Lucia Widow at a temporary pool of water in Giyani or a beautiful Streamer Tail moth in Tzaneen (see below). Just by chance, I was talking on my cellphone outside our offices in Tzaneen when the Streamer Tailed moth fluttered by. In a few slick, ninja-like moves, I managed to catch the moth, take a few photographs and release it again – much to the amusement of my work colleagues. I have yet to find another of these gorgeous moths. It is elusive species such as this which I hope to still encounter on my journey.
The Streamer tail moth caught outside my place of work in Tzaneen.
I try to reach 1000 records per year for the Virtual Museum. This is the first year that I have struggled to reach this target. This is largely due to the sudden drop in Lepidoptera numbers, a phenomenon observed across most parts of South Africa, in the summer of 2017/18..
BDI: How do you react to the statement that “Being a citizen scientist is good for my health, both physical and mental!”?
Citizen science really inspires a person to get out there and explore the environment – thus pursuing healthy activities. Within my line of work, one unfortunately often has to deal a lot with the darker side of life. In my spare time, spending time in the environment looking for new records which can contribute towards citizen science projects, and the excitement of finding something new, really helps me to unwind and de-stress. This helps me to cope in what can be a very hectic, demanding environment.
“Disaster management” is the business of turning chaos into order. There are no dull moments! Something is always demanding attention. Citizen science helps me to unwind and de-stress after a hectic week or event.
What do you see as the role which citizen science plays in biodiversity conservation? What is the link?
There is most certainly a link between citizen science and the conservation of biodiversity. Without the knowledge which is being generated by citizen science, I think it would be far more difficult to really monitor biodiversity. At the same time, there is a great deal of awareness which is being raised which contributes towards conservation and the promotion of biodiversity. In a rapidly changing world under climate change, the massive number of records generated through citizen science can broaden our understanding of how the distribution of different species is changing, and ultimately how this may impact on the human environment as well. We cannot live on this planet and think we are in no way connected with the natural environment. We depend on the environment, and if the environment suffers, so do we. Citizen science can make significant contributions by reaching a broad range of people, who in turn inspire and teach others this very basic concept.
Systematic atlasing in Hessequa – moving from mapping to monitoring
Johan A van Rooyen*
U3A Stilbaai – Bird Group
Introduction
Les Underhill and Michael Brooks in their 2016 paper about the progress of the Second Southern Africa Bird Atlas Project (SABAP2), mention the Stilbaai Bird Club in the paragraph “Personal, club and group challenges and targets”. In this paper, I document the targets set and achieved by the atlasers of the Stilbaai Bird Group over the period 1 October 2014 to 30 November 2017 (Underhill & BrooKs 2016).
In September 2014, the Stilbaai Bird Club was introduced to the SABAP2 protocol (fully described in Underhill 2016). Some members started atlasing in October 2014. The atlasing of the Stilbaai atlasers is coordinated by the author of this paper. In 2016 the bird club was integrated into the U3A: Stilbaai as a Bird Group.
The Stilbaai atlasers started off as a group of 10 atlasers of which eight are husband and wife teams and in 2017 they were joined by two more atlasers. All of them are retired and live in Stilbaai, except for one couple who reside in George.
Because Stilbaai falls in the Hessequa Municipality, it was decided to atlas the pentads that cover the municipal area. The area is shown in Figure 1 and lies more or less between the Breede River in the west and the Gouritz River in the east, and the Langeberg Mountain in the north and the ocean in the south. A few pentads without access roads in the Langeberg Mountain were excluded, as well as two pentads in the south on private land where access could not be arranged. The resultant “Hessequa Atlas Area” comprises 75 pentads. For ease of reference the pentads were all given names of farms or towns or other prominent features over and above the standard reference number and those names are used by all the atlasers in the group.
Fig 1. Map of Hessequa atlas area
This is a large area of about 110 x 55 km with the farthest pentads between 90 and 100 km from Stilbaai. This area forms the eastern half of the Overberg and is almost entirely rural. Most of the area is agricultural, where the main crop is wheat. In the south, much of the coastal fringe is natural vegetation. The northern pentads lie along the Langeberg mountain range.
The objectives set by the group for each of the years evolved over time, but it was realised right from the outset that it must accommodate the two broad objectives of SABAP2. That is to improve the information for mapping purposes, as well as to move towards monitoring as a longer term goal. The objectives and achievements for each year are given in more detail in the rest of the paper.
It is important to realise that it is not only the Stilbaai atlasers who are doing SABAP2 fieldwork in this area. The coastal towns of Witsand, Stilbaai/Jongensfontein and Gouritsmond are popular holiday destinations and have attracted a fair share of visiting atlasers. The same applies to the Grootvadersbosch Nature Reserve and the very popular Voëlvlei near Gouritsmond. A number of visiting atlasers also contributed significantly in the rest of the area.
Coordination of atlasing in Hessequa
Atlasing systematically in Hessequa is coordinated by using a semi-automated spreadsheet system that incorporates the specific targets for any given year. The Stilbaai atlasers are issued with a priority list of pentads by email at the start of each month from which each atlaser then chooses the pentads they want to visit. Each atlaser informs the others of his/her choice by using “reply to all” to prevent duplication. The list is also updated weekly to show the choices already made and pentads completed.
The spreadsheet is updated regularly by keeping track of atlas cards that are submitted for the Hessequa area. A “Pentad Group” covering the Hessequa area was created on the SABAP2 website (Figure 2) and this shows information only for the 75 pentads. This simplifies the monitoring of submitted cards tremendously. Many cards are submitted by visitors to the area and this is the most efficient way of keeping track of those cards as well. The information from the website is copied to the spreadsheet to mirror the website and allows for calculations to compare cards submitted to the targets that were set.
Fig 2. Hessequa pentad group on SABAP2 website
Base conditions
In the remainder of this paper the pentads of the Hessequa atlas area are shown schematically and the number of completed full protocol cards for each pentad is shown in the same colours as on the SABAP2 website. The legend is given in Figure 3 for easy reference.
Fig 3. Legend indicating number of full protocol cards per pentad
In each cell in the following figures the top line is the pentad number, the second line the Stilbaai atlasers’ name for the pentad and the third line the number of full protocol cards.
The cumulative coverage of the Hessequa area from the start of SABAP2 in July 2007 up to the end of September 2014 is shown in Figure 4. At the start of this initiative, only 15 of the 75 pentads had one full protocol card and 27 pentads had reached the foundational coverage of four or more cards (Figure 4).
Fig 4. Cumulative full protocol cards up to 30 September 2014, at the start of systematic atlas fieldwork in the Hessequa
Atlasing in 2015
The Stilbaai atlasers started slowly in the last quarter of 2014 but firm targets were set annually from the beginning of 2015.
The first target for 2015 was to increase the coverage for mapping purposes and the specific aim was to turn Hessequa green, that is a minimum of four cards per pentad. It was, however, also agreed by the atlasers that more than that could be achieved and two quarter degree squares, one around Stilbaai (3421AD) and the one just north of it (3421AB) were selected for more intense atlasing. This area is called the Stilbaai core area and the aim was to turn it dark green with seven cards per pentad.
The second target for 2015 was already focusing on the monitoring objective of SABAP2 and it was decided that each pentad will be atlased at least once regardless of the number of cards already submitted.
The 2015 coverage that was achieved is shown in Figure 5. A total of 195 full protocol cards were submitted, 152 by the Stilbaai atlasers and 43 by visitors. The monitoring target of a minimum of one card per pentad was thus achieved. The cumulative coverage by 31 December 2015 is given in Figure 6.
Fig 5. Full protocol cards submitted during 2015, the first year of systematic atlas fieldwork in the HessequaFig 6. Cumulative full protocol cards up to 31 December 2015 at the end of the first year of systematic atlas fieldwork in the Hessequa
The target of “turning Hessequa green” was, except for three pentads, achieved by the end of 2015 and the coverage of the Stilbaai core area was increased to seven cards or better (Figure 6).
Atlasing in 2016
The monitoring target was set slightly higher for 2016 to a minimum of one card per pentad for the whole area, except for the Stilbaai core area where it was set as a minimum of 4 cards. It was also decided to atlas the two home pentads around Stilbaai (3420_2120 and 3420_2125) at least once a month.
The mapping target was to get the Stilbaai core area to 11 cards (light blue). A longer term mapping target was to get the rest of Hessequa to a minimum of 7 cards (dark green) by the end of 2017. In order to spread the work load between 2016 and 2017 some pentads would then have to be atlased twice during 2016.
During 2015 no attention was paid to ensure an equal distribution of cards over the months of the year for each pentad. For 2016 and subsequent years, it was decided to aim for as equal a distribution as practically possible and pentads were prioritised on a monthly basis to achieve this target. The result of what was achieved to 30 November 2017 is discussed later on.
The 2016 coverage is shown in Figure 7. A total of 277 full protocol cards were submitted, 175 by Stilbaai atlasers and 102 by visitors.
The monitoring targets were met except for pentad 3355_2050. During a data quality control exercise during 2017 it was found that a number of cards that were submitted as full protocol cards did not cover the required minimum of two hours atlasing. They were subsequently changed to ad hoc cards and the one full protocol card that was submitted for 3355_2050 for 2016 was changed to an ad hoc card. Quality control is now done on a continuous basis in order to prevent this from occurring again.
The cards submitted for pentads 3420_2120 and 3415_2120 were much higher than expected due to the out-of-range Red-necked Buzzard that caused a huge influx of “twitchers” who also submitted full protocol cards. But as Underhill & Brooks (2016) stated: “There is no pentad for which SABAP2 has”enough” checklists.”
The cumulative coverage by 31 December 2016 is given in Figure 8. All pentads had by this stage reached the minimum mapping requirement of 4 cards, the Stilbaai core area the 11 card target and some of the others had by now reached the 7 card (dark green) stage.
Fig 7. Full protocol cards submitted during 2016, the second year of systematic atlas fieldwork in the HessequaFig 8. Cumulative full protocol cards up to 31 December 2016, at the end of the second year of systematic atlas fieldwork in the Hessequa
Atlasing in 2017
After a discussion with Prof Les Underhill late in 2016, it was decided to increase the monitoring target for Hessequa. The target for 2017 was set for a minimum of two full protocol cards per pentad for the whole area.
This target was already achieved by 30 November 2017 and is shown in Figure 9. A total of 267 full protocol cards was submitted, 212 by Stilbaai atlasers and 55 by visitors.
Fig 9. Full protocol cards submitted during 2017, the third year of systematic atlas fieldwork in the Hessequa
The cumulative coverage up to 30 November 2017 is shown in Figure 10 and the whole of Hessequa was by this date dark green (seven or more cards).
Fig 10. Cumulative full protocol cards up to 30 November 2017, at the end of the third year of systematic atlas fieldwork in the Hessequa
Targets for 2018
In further communication with Prof Les Underhill during 2017, it was recommended that an even more ambitious monitoring target be set for 2018. That is to again do two full protocol cards per pentad, but to distribute these cards evenly over the four seasons of the year.
The four seasons are defined as follows:
Summer – December, January and February
Autumn – March, April and May
Winter – June, July and August
Spring – September, October and November.
It is this definition that forced the “shortening” of the 2017 atlas year to end by 30 November in order to start with the new programme on 1 December 2018.
The Stilbaai atlasers, however, decided to increase the 2018 target by aiming to do four cards per pentad (one card per season) for the Stilbaai core area. The two home pentads will again be atlased at least once per month.
The target for 2018 is shown schematically in Figure 11. In this figure each pentad has four rows, one for each season starting with summer at the top.
Fig 11. Atlas targets in the Hessequa for 2018, showing planned seasonal distribution of effort
Figure 11 shows that the 2018 target for the Stilbaai core area is a card in every season and three cards per season for the Stilbaai home pentads (continuing with the one card per month objective). In the rest of Hessequa the target is for each pentad to get a card for summer and winter while the adjacent pentads will get cards for autumn and spring in a chessboard pattern. If this is achieved, the pentads will be switched around in 2019 to cover all four seasons in a two year cycle. Two exceptions to this rule can be seen in the northeastern and northwestern corners of Hessequa where access problems make it a logistical imperative for two adjacent pentads be done at the same time.
If the above targets are achieved, the Stilbaai core area will be turned dark blue (16 cards) during 2018 and the rest of the Hessequa area light blue (11 cards) which will provide excellent information for the objective of mapping bird distributions.
Monthly distribution of full protocol cards
Since the start of 2016 the Stilbaai atlasers have aimed to spread the atlas cards as evenly as practically possible over the months of the year. The monthly distribution of the cards as on 30 November 2017 is shown in tabular form in Figures 12, 13 and 14. The colours used in the monthly columns are the standard colours used throughout this paper.
Fig 12. Monthly distribution of cards in the Hessequa achieved up to 30 November 2017 (Pentads 1-25)Fig 13. Monthly distribution of cards in the Hessequa achieved up to 30 November 2017 (Pentads 26-50)Fig 14. Monthly distribution of cards in the Hessequa achieved up to 30 November 2017 (Pentads 61-75)
It can be seen from Figure 12, 13 and 14 that the distribution of the cards over the months of the year is very good, except for the pentads where a large number of cards have been done by visitors. That is perfectly understandable as they often visit the area during the same time of the year and those cards obviously contribute greatly towards the detailed understanding of species distribution during those months.
Conclusions
From 1 October 2014 to 30 November 2017, the Stilbaai atlasers, with a steady stream of cards from visitors, improved the foundational coverage of the Hessequa area. Initially 20% of pentads had one card and about a third of pentads had four cards (Figure 4). At the end of this period, the coverage depth was seven cards or more for every one of the 75 pentads of the Hessequa area and 11 or more cards for the Stilbaai core area.
This was achieved while simultaneously achieving monitoring targets of at least one card per pentad for 2015 and 2016 and two cards per pentad for 2017. An excellent distribution of full protocol cards over the months of the year was also achieved. This will be taken forward into the future.
With the foundational coverage on a sound footing, the Stilbaai atlasers will focus on more intensive monitoring in future and will strive for an even distribution of full protocol cards over the seasons of each year. This paper describes a good example of what can be achieved by a group of birders working together with a systematic strategy towards atlasing goals.
Acknowledgements
We are greatly indebted to the 146 visiting atlasers who built the base from 2007 to 2014 and who are still contributing large numbers of cards every year. The following atlasers contributed 10 or more cards to the Hessequa area and deserve a special mention: Pieter de Waal La Grange (73), Don Reid (34), Josef van Wyngaard (20), Pat Nurse (19), Brian Vanderwalt (18), Stefan Theron (17), John Carter (16), Tineke Malan (16), Garth Shaw (16), Adrius Rabie (14), Alan Collett (10), Maria Kemp (10) and Hans Jurie Linde (10).
The Stilbaai atlasers are a special group of people who took to atlasing with great enthusiasm and have been willing to do their atlasing in a coordinated way since October 2014. Without their unwavering support, this would not have been possible. They are Patrick and Alice Duddy, Ben and Joekie Jovner, Phil and Esther Scheffer, Hennie and Gerda Smit, Menno Stenvert and Krysia Solman, Terry and Maria Terblanche, Annelie van Lill, Johan and Estelle van Rooyen, Rita van Rooyen, John and Jeannie Willemse, Dirk Bosman and Derick Oosthuizen.
References
Underhill LG 2016. The fundamentals of the SABAP2 protocol. Biodiversity Observations 7.42:1-12.
Underhill LG, Brooks M 2016. SABAP2 after nine years, mid 2007-mid 2016: coverage progress and priorities for the Second Southern African Bird Atlas Project. Biodiversity Observations 7.37:1-17