Find the Plaintive Rain Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Brevicepitidae
Identification
The female Plaintive Rain Frog attains a length of 53mm (males are smaller than females). The body is stout with short legs and relatively small eyes. The colouration of the upper sides is variable but is most often tan-brown to dark brown with black markings. Specimens from forested places can be mostly black, sometimes with paler spots. Both the upper and lower parts are granular.
The natural habitats of the Plaintive Rain Frog are indigenous forest and the adjacent grassland along the eastern escarpment. Along the great escarpment, the Plaintive Rain Frog occurs in Moist Upland Grassland, North-eastern Mountain Grassland, Afro Mountain Grassland, and adjacent Afromontane Forest. Along the Eastern Cape and KwaZulu-Natal coast, it inhabits Coastal Bushveld/Grassland, Coastal Forest, Sand Forest, and Dune Thicket. At Pennington in KwaZulu-Natal (3030BC), males were heard calling in dense, matted grass, five metres from the high-water mark (Minter 1998). Between the coast and the escarpment in KwaZulu-Natal, it is found in Short Mistbelt Grassland. At Eshowe, calling males were collected in suburban gardens (Minter 1998).
Coastal forest habitat. Mtunzini, KwaZulu-Natal hoto by Ryan Tippett
Factors that are common to these vegetation types, and may limit the distribution of this species, are annual rainfall in excess of 700 mm and well-shaded soil, which result in higher soil moisture levels over relatively longer periods than is the case in drier, more open habitats.
Behaviour
During dry periods, the Plaintive Rain Frog remains below the surface. Minter (1998) followed a shallow burrow for 3 m until it dipped down and disappeared under a large rock, and found deep, vertical burrows near the call sites of several males. Specimens have also been collected under rocks (Jacobsen 1989).
Plaintive Rain FrogBreviceps verrucosus Baziya forest, Eastern Cape Photo by Luke Kemp
The breeding period of the Plaintive Rain Frog is relatively short, commencing with the first spring rains in August or early September and seldom extending beyond mid-November. Choruses develop rapidly in rain, drizzle or heavy mist and continue unabated, day and night, as long as the wet weather lasts. Males are prompted to vocalize by the calls of their immediate neighbours, and in this way bouts of calling spread through the population in waves, as in the Bushveld Rain Frog B.adspersus. The Plaintive Rain Frog is easily prompted to call by a tape recording or human imitation of the call, even when no other calling is taking place (pers. obs.). Calling is inhibited by wind and low temperatures (Minter 1998).
The call site is usually a well-concealed, shallow depression under dense, matted grass or leaf litter, but males may call at the surface, or from elevated positions on grass tussocks, up to 40 cm from the ground (Poynton and Pritchard 1976; Minter 1998). Amplexus, oviposition and development have not been recorded.
Food items include amphipods, isopods, juvenile myriapods, caterpillars, beetles and ants (Poynton and Pritchard 1976).
Status and Conservation
The Plaintive Rain Frog is widely distributed in a variety of habitats and does not appear to be at risk. It occurs in a number of provincial nature reserves and in the Golden Gate Highlands National Park (Bates 1995).
Plaintive Rain Frog Breviceps verrucosus Mariepskop, Limpopo Photo by Luke Verburgt
Distribution
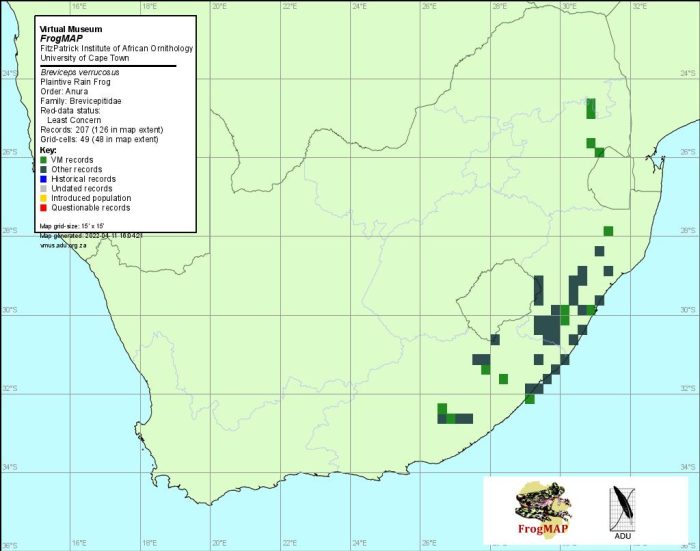
The Plaintive Rain Frog is endemic to South Africa, Lesotho, and eSwatini (Swaziland). It occurs along the Great Escarpment, from the Nico Malan Pass (3226DA) in Eastern Cape Province, northward through Lesotho, KwaZulu-Natal and Swaziland, to the Blyde River Canyon Nature Reserve (2430DB) in Mpumalanga. A single specimen was collected on the border of Lesotho, in eastern Free State (Bates 1995). The species descends into moist grassland and forest east of the escarpment, reaching the coast at Coffee Bay (3129CD), and extending along the coast as far north as Empangeni (2831DD). The most southerly record is based on a museum specimen from Zuurberg (3325BC).
The great variation in colour and markings present in different populations of this species is attributed to crypsis. For example, individuals living in forest and along the forest fringe at Graskop (2430DD) are black with fine, cream speckles, while those living on light sandy soils at Eshowe are uniformly light brown (Minter 1998). Populations from grassland habitats in Eastern Cape Province exhibit a varying degree of light brown mottling on a dark brown background, and were previously referred to a separate subspecies, B. v. tympanifer. However, similar marking are seen on specimens from grassland habitats in KwaZulu-Natal and Mpumalanga, while a population sampled at the grassland/forest interface at Wesa (3029DA) contains entirely melanistic as well as mottled individuals (Lambiris 1989a; Minter 1998). Therefore, B. v. tympanifer does not represent a valid subspecies, but rather a cryptic colour variation in grassland (Minter 1998). These various populations show no differences in advertisement-call structure, which confirms that they are conspecific (Minter 1998).
Distribution of Breviceps verrucosus. Taken from the FrogMAP database. April 2022.
Breviceps maculatus, described from montane grassland in the KwaZulu-Natal Drakensberg (FitzSimons 1947), has been recognized by some authors as a distinct species (Poynton 1964; Lambiris 1989a, 1991) on the basis of dorsal markings and larynx structure, while others have placed maculatus in the synonymy of B.verrucosus because of the similarity of their advertisement calls (Passmore and Carruthers 1979, 1995; Bates 1995; A. Channing pers. comm.; pers. obs.). B. maculatus is not treated as a separate taxon in this atlas, but an analysis of a larger sample of specimens and calls is required in order to validate this viewpoint.
The atlas data are reliable, but incomplete. The taxonomic status and distribution of populations in Lesotho, and along the escarpment in Mpumalanga, and possibly Limpopo Province, warrant further investigation (see B. sylvestris species account).
Further Resources
The use of photographs by Cornelia Rautenbach, Luke Kemp, and Luke Verburgt is acknowledged.
Recommended citation format: Minter, LR; Tippett, RM. (2025). Plaintive Rain FrogBreviceps verrucosus. Biodiversity and Development Institute, Cape Town. Available online at https://thebdi.org/2021/12/15/plaintive-rain-frog-breviceps-verrucosus/
This species text has been updated and expanded from the text in the 2004 frog atlas: Minter, LR. (2004). Plaintive Rain FrogBreviceps verrucosus. In Minter LR et al 2004.
References:
Minter, LR; Burger, M; Harrison, JA; Braack, HH; Bishop, PJ; Kloepfer, D. (Editors). (2004). Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
Carruthers, V; du Preez, L. (2017). Frogs of southern Africa: A Complete Guide. Struik Nature, Cape Town.
Channing, A. (2001) Amphibians of Central and Southern Africa. Protea Book House, Pretoria
Claus, B; Claus, R. (2002). Common Amphibians and Reptiles of Botswana. Gamsberg Macmillan, Windhoek.
View the above photo record (by Nick Evans) in FrogMAPhere.
Find the Plain Stream Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pyxicephalidae
PLAIN STREAM FROG – Strongylopus wageri
Wager, 1961
Identification
S. wageri reaches 50 mm in body length. The dorsal colouration varies from uniform light brown or yellow-green to brick red, and is slightly paler on the snout. A distinct, dark band passes from the nostril through the eye to the shoulder and then continues along the flank. Faint barring on the legs and a narrow, pale vertebral stripe may be present (Wager 1965; Lambiris 1989a).
S. wageri may be distinguished from S. grayii, which sometimes has a similar dorsal colour pattern, by the extent of the webbing on the longest toe: in S. wageri no more than three phalanges are free of webbing, whereas in S. grayii, four or occasionally three phalanges are free of webbing (Poynton 1964; Passmore and Carruthers 1995).
The advertisement call is a short cackle c.500 ms in duration, repeated at irregular intervals. A call consists of a variable number of short notes, each about 50 ms in duration (Passmore and Carruthers 1995; Channing 2001).
Wager (1986) noted that tadpoles from Weza, Giant’s Castle and Mont-aux-Sources were identical, but Lambiris (1989a) found that adults from both Giant’s Castle Game Reserve and Entumeni Nature Reserve differ from the type series (Weza) with regard to webbing and colour pattern and suggested that they may represent an undescribed sibling species. The taxonomic status of the various isolated populations that do not form part of the main KwaZulu-Natal Drakensberg population, should also be examined as they may be in the process of speciation.
Habitat
At lower altitudes, S. wageri inhabits mistbelt forest, while at high altitudes up to 2000 m, it occurs in montane grassland. Adults may be found amongst vegetation or stones on the banks of clear streams and pools. Breeding takes place in clear, cold, fast-flowing streams and their pools.
Behaviour
Adults are active in the late afternoon and at night. When disturbed they jump into the water and remain motionless in the mud at the bottom, apparently relying on camouflage to avoid detection.
S. wageri breeds January–May, but mainly in autumn when water levels have stabilized (Channing 1979). Males call from concealed or partly concealed positions along the edges of streams or pools and sometimes under water (Lambiris 1989a).
In the laboratory, spawning was recorded at about 03:00. Clutches of 12–250 eggs are firmly attached to twigs or blades of grass dangling or submerged in the water, or to submerged rocks. The eggs measure 2.8 mm in diameter and are enclosed in a stiff, jelly-like capsule with a diameter of 5 mm, that swells to 7 mm after a few days. Eggs hatch after 8–9 days (Wager 1986; Channing 1979).
The tadpoles lie in the algae and mud at the bottom of the pools, moving slowly, but darting around rapidly if disturbed. They are often found in sunny positions in shallow water (Channing 1998). Tadpoles grow slowly in the cold of winter, and metamorphosis is completed after 5–9 months.
Adult S. wageri prey on terrestrial arthropods and gastropods (Channing 1979), as well as a variety of aquatic and semi-aquatic insects (Lambiris 1988e). In turn, they are probably eaten by a variety of vertebrates such as birds, snakes, otters and other frogs. Channing (1979) observed a crab climbing onto vegetation to devour the eggs of this species.
Status and Conservation
Status
S. wageri is placed in the category Near Threatened (Harrison et al. 2001; this publication) because of its small extent of occurrence and area of occupancy, fragmented distribution, and an existing and projected decline in the extent and quality of its habitat over a large part of its range. The species was not placed in a threatened category because it is relatively well protected in the reserves along the KwaZulu-Natal Drakensberg. It should be noted that S. wageri is heavily dependent on the continued protection of its habitat in these areas.
S. wageri is recorded from several conservation areas in KwaZulu-Natal, namely Ukahlamba Drakensberg Mountain Park, as well as Entumeni Nature Reserve, Ntsikeni Nature Reserve, Qudeni State Forest, Ngele Forest Nature Reserve and Ngome Forest Reserve.
Threats
In the past, S. wageri was probably more widespread on the Drakensberg escarpment in Mpumalanga, than at present. Much upland habitat in these areas has been destroyed by intensive afforestation with alien trees, and this is likely to have affected local populations of S.wageri. Channing (1979) noted that the destruction of indigenous forests has probably also resulted in loss of habitat to this species in KwaZulu-Natal.
Habitat loss to S. wageri is estimated to be >20% over the past 50 years, with a similar rate of decline in population numbers due to afforestation, chemical pollution associated with sylviculture, alien-plant invasions and the introduction of trout into streams (Harrison et al. 2001).
Recommended conservation actions
Extensive distribution surveys and monitoring of known populations are recommended. Habitat management, including the identification of limiting factors, is needed for effective conservation of this species (Harrison et al. 2001). It is especially important to determine whether S. wageri is able to survive in streams that run through plantations.
Distribution
S. wageri is endemic to South Africa. It has not been recorded at or near the summit of the Drakensberg in Lesotho (Bates and Haacke 2003). Distribution records are sparse and widely scattered. From the type locality on the Ngele Mountains near Weza Forest (3029DA) in KwaZulu-Natal, the species ranges northward along the escarpment and foothills of the Drakensberg mountain range. Isolated populations exist further to the east in KwaZulu-Natal, at Qudeni Forest (2830DB), Entumeni Nature Reserve (2831CD) and Ngome Forest Reserve (2731CD), and along the southern boundary of Mpumalanga Province.
Acocks (1975) suggested that in the recent past (A.D. 1400) KwaZulu-Natal was covered by forest and scrubforest, with grassveld on the summit of the Drakensberg. If so, this may explain the present disjunct distribution of this species in patches of forest that were once connected (Channing 1979).
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Wager se langtoonpadda (Afrikaans)
Recommended citation format for this species text:
Bates MF, Tippett RM. Plain Stream Frog Strongylopus wageri. BDI, Cape Town. Available online at http://thebdi.org/2021/12/15/plain-stream-frog-strongylopus-wageri/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Bates MF 2004 Strongylopus wageri Plain Stream Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Felicity Grundlingh) in FrogMAPhere.
Find the Cape Platanna in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pipidae
CAPE PLATANNA – Xenopus gilli
Rose & Hewitt, 1927
Identification
X. gilli is a typical member of the genus Xenopus. The upper body is light to yellow-brown with elongated, dark brown patches, sometimes paired, that begin between the eyes and extend backwards, breaking up into smaller patches on the lower back and upper surfaces of the hind limbs. The underside usually has clear blackish and yellow mottling, but this may be pale and indistinct in some individuals, especially in populations near Kleinmond (3419AC; J.A. Harrison in litt.).
X. gilli can be distinguished from X. laevis by the following morphological features: its smaller size (<60 mm in body length); a narrower, more acutely pointed head; the absence of a subocular tentacle (present but inconspicuous in laevis); a poorly developed inner metatarsal tubercle (a distinct ridge in laevis); and less conspicuous lateral line sense organs (Poynton 1964; Picker and de Villiers 1988; Passmore and Carruthers, 1995; Kobel et al. 1996; Channing 2001).
The advertisement call consists of a series of short, rapidly pulsed, metallic buzzes emitted under water at a rate of about two per second (for further details of call structure see Passmore and Carruthers 1995; Picker et al. 1996; Channing 2001).
Habitat
X. gilli inhabits blackwater wetlands in low-lying coastal areas. These are permanent and seasonal seepages, marshes, ponds, pans, vleis and coastal lakelets, in a variety of fynbos vegetation types and, in places, a mixture of fynbos and dune thicket. The vegetation types include mostly Mountain Fynbos, Sand Plain Fynbos (on the Cape Flats), or Mountain Fynbos mixed with either Limestone Fynbos or Dune Thicket. The substrate has a predominantly sandy base and varies, depending on the humic content, from white or grey to a dark brown or blackish soil.
The water is humic and dark in colour, low in nutrients, high in dissolved solids, and typically has a low pH (minimum 3.4; Picker 1985). It has been demonstrated that the tadpoles of X. gilli can tolerate pH as low as 3.6, whereas X. laevis tadpoles have a reduced rate of survival below pH 5–6. This accounts for the observed habitat segregation between these species on the Cape Peninsula where X. gilli occurs in acidic blackwater seepages and ponds, while X. laevis prefers clear water bodies with elevated pH, for example, artificial impoundments (Picker et al. 1996). Disturbances of X. gilli water bodies which alter the humic content and nutrient levels, cause an increase in pH levels and often result in the colonisation of these water bodies by X. laevis, providing an opportunity for hybridization between the species (Simmonds 1985; Picker et al. 1996).
While X. gilli avoids habitats that have been disturbed by urban development or agriculture, or that contain invasive plants and animals (Picker 1985; Picker and De Villiers 1989), it is interesting to note that healthy populations of this frog inhabit certain seepages that were excavated to form waterholes in the Cape of Good Hope area of the Cape Peninsula National Park (CPNP). However, no other habitat disturbances or threats are evident in these pools, the water quality meets the necessary requirements, and consequently X. gilli populations have flourished in them for well over two decades.
Behaviour
When its wetland habitat dries up during the summer months, X. gilli survives by aestivating below the surface. Rau (1978) found several specimens encapsulated in the mud of dried-up vleis on the Cape Flats, and Rose (1962: 33) found one individual at a depth of 15 cm amongst the roots of a large “weed” growing on the site of a small dried-up vlei. During the rainy season, overland migrations between ponds have been observed in Cape Peninsula National Park (Picker 1985).
Breeding commences during the wet winter months (July), and continues until late October (Rau 1978). Three to four hundred dark brown eggs are laid over a period of a day, each surrounded by a jelly capsule 1.3 mm in diameter (Channing 2001). The nektonic tadpoles feed on phytoplankton in the water and complete their metamorphosis by the end of summer. Rau (1978) recorded spawning activity over a four-month period and found metamorphosis to take about 120 days. The breeding season of X. gilli overlaps that of X. laevis, which increases the opportunity for hybridization (Rau 1978; Picker et al. 1996).
Adult frogs feed on living and dead animal material in their wetland habitat, including aquatic invertebrates and the eggs, tadpoles and smaller frogs of their own kind and other species (Picker and De Villiers 1988). Predation of the immature stages of X. gilli by the larger X. laevis is presumably intense in disturbed habitats that have been invaded by the latter. Other predators include herons, cormorants and water mongoose.
Status and Conservation
Status
Development and general habitat degradation have severely impacted on the extent of occurrence and area of occupancy of this species, resulting in a loss of more than 50% of its habitat, and severe fragmentation of its populations (Harrison et al. 2001). This is particularly serious on the Cape Flats and adjoining Cape Peninsula where extensive urban development has taken place. By now, X. gilli is possibly extinct in both of these areas except for populations in the Cape Peninsula National Park. Similar threats are escalating between Rooiels and Kleinmond on the south coast, leading to further habitat loss and fragmentation.
About 70% of all currently known X. gilli habitat is situated in the Cape Peninsula National Park. This is the stronghold of the species with healthy populations in the Cape of Good Hope area. The populations at some sites in this area have been found to vary from 121–591 frogs (Picker and De Villiers 1989; Picker et al. 1996). It is nevertheless of concern that X. laevis and/or hybrids have been reported from most of the wetlands in this area, and the situation needs to be monitored. It is also of concern that the only X. gilli habitat protected within a conservation area, besides Cape Peninsula National Park and Greater Betty’s Bay Nature Reserve, is a small remnant of habitat in Agulhas National Park.
X. gilli was included in the first South African Red Data book for amphibians, in the Rare category (McLachlan 1978). In the revision (Branch 1988), it was classified Endangered. Endangered status was retained in Harrison et al. (2001), based on an extent of occurrence <5000 km2, an area of occupancy <500 km2, a severely fragmented habitat, continuing decline in the extent of occurrence, area of occupancy, extent and quality of habitat and the number of locations/subpopulations and mature individuals. The species is legally protected by Nature Conservation Ordinance 19 of 1974, but is not listed by CITES.
Threats
Loss of habitat and habitat fragmentation pose the most serious threats to the survival of X. gilli. In particular, urban development and agriculture have resulted in the filling in and drainage of its wetland habitat or have led to the pollution and eutrophication of breeding sites. Furthermore, the building of artificial reservoirs and irrigation systems has enabled the highly adaptable X.laevis to invade areas from which it was previously excluded, including disturbed blackwater wetlands containing populations of X. gilli. Predation by X. laevis on the eggs, tadpoles and froglets of X. gilli represents a further threat to the survival of X. gilli.
Urban expansion and human activities also accelerate the spread of invasive alien vegetation. At some localities, indigenous fynbos vegetation has been replaced by stands of exotic trees (Port Jackson Willow Acacia saligna and Rooikrans A. cyclops). This alters the water chemistry and results in unsuitable habitat for X. gilli. Introduced predatory fish may pose an additional threat, particularly in some of the larger, permanent wetlands.
The invasion of disturbed X. gilli habitat by X. laevis, and the subsequent hybridization of these two species, has been well documented (Rau 1978; Picker 1985; Simmonds 1985; Picker and De Villiers 1989; Picker et al. 1996; Evans et al. 1998). Hybridization threatens the gene pool of the smaller, less numerous X. gilli, through potential genetic swamping of populations. This threat is present throughout the distribution area of X. gilli (Picker et al. 1996) but appears not to be as serious as was previously thought (Evans et al. 1998).
Recommended conservation actions
The distribution and conservation status of X. gilli is monitored by the Western Cape Nature Conservation Board (WCNCB) as part of a threatened species monitoring programme (De Villiers 1997). Healthy populations in the Cape of Good Hope area were monitored mainly by the Zoology Department of the University of Cape Town, but monitoring ceased in 2000. It will be continued by the CPNP in conjunction with the WCNCB. Besides the monitoring of known localities, additional survey work is to be conducted in surrounding areas, including montane habitats.
Although X. gilli populations on the Cape Peninsula are well protected, it is important that other viable populations be included in statutory conservation areas and managed appropriately. This was strongly recommended by Evans et al. (1997), who found that populations to the east of False Bay showed significant genetic differences from the Cape Peninsula populations, although these were not considered to be taxonomically significant. In particular, they indicated that “protective measures within X. gilli habitat near Kleinmond would conserve much of the genetic diversity seen in this species”.
The main management activity is control of alien vegetation. This is undertaken on the Cape Peninsula by CPNP, in the Kleinmond area by the WCNCB, and in the Betty’s Bay area by the Overstrand Municipality. Alien vegetation clearing programmes are now underway in the Gansbaai to Agulhas area, but they need to be intensified in X. gilli habitat.
In 1985, a Cape Platanna Conservation Committee built a precast wall around Geps Dam, one of the pools in the Cape of Good Hope area, to protect its X. gilli population from contamination with X. laevis (Picker and de Villiers 1989). After the wall was built, X. laevis and hybrids were removed from the water body. Holes were regularly found under the wall, but no X. laevis or hybrids were observed in the water body when it was last examined in 2000. In fact, X. laevis and hybrids are apparently on the decline in at least this section of the Cape of Good Hope area (M.D. Picker pers. comm.). Although the situation requires continued monitoring, it seems that the wall now serves little purpose and might safely be removed.
In 1988, the Cape Platanna Conservation Committee translocated 154 juvenile X. gilli from the Cape of Good Hope area to four blackwater pools in the Silvermine nature area, both areas falling within the present CPNP. Although a survey produced no sign of X. gilli in the Silvermine nature area shortly before the translocation, about 16 specimens were collected in 1926 from somewhere in and next to the “Sylvermyn River” which drains this area (Rose and Hewitt 1927). The main reason for the translocation was to establish a separate breeding colony of X. gilli on the Cape Peninsula that would perhaps be free of the X. laevis threat. It would appear that this experiment has had some success: from one to six adults have been seen in one of the pools on about four occasions during the 10-year period following the translocation. Further recommendations are being formulated in this regard, and monitoring work is to continue.
Distribution
This species is endemic to the winter rainfall region of the Western Cape, generally occurring in relatively low-lying areas (10–140 m a.s.l.) within 10 km of the coastline. Its distribution is correlated with the presence of nearby mountain ranges and an annual rainfall exceeding 500 mm p.a. Confirmed records span a distance of about 160 km, from the Cape Peninsula southeastward towards the Agulhas district (Picker and de Villiers 1988, 1989).
For about 40 years following its description in 1927, the species was known only from the Cape Peninsula and adjoining Cape Flats (3418AB, BA; 3318CD, DC). However, in the late 1960s it was discovered further south on the Cape Peninsula in the Cape of Good Hope Nature Reserve (3418AD), and in the period from 1973 to 1988 it was collected along the coastal forelands to the southeast of the Cape Peninsula and Cape Flats, at Hangklip and Betty’s Bay (3418BD), Kleinmond (3419AC), midway between Gansbaai and Agulhas (3419DA), and northwest of Agulhas at the eastern base of Soetanysberg (3419DB, DD).
Two inland records from Nieuwoudtville (3119AC) in 1898 and Citrusdal (3219CA) dating from about 1937 (Rau 1978), have not been subsequently confirmed despite several field trips to these areas. The validity of these records is in doubt and they are excluded from the map.
During the 1980s a study of the distribution and habitat requirements of X. gilli was undertaken to assess its future survival prospects. The study showed that, although the species still occurred in at least 24 localities in seven quarter-degree grid cells, habitat destruction and degradation had led to extinction at 60% of its known localities (Picker and De Villiers 1989).
Thus,X. gilli has been recorded from a total of 10 quarter-degree grid cells (excluding the two inland localities), but since 1995 it has been found in only five of these cells (3418AB, AD, BD; 3419AC, DA). The atlas data are reliable.
Distribution of Xenopus gilli. Taken from the FrogMAP database as at November 2021.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gill’s Platanna (Alternative Common Name); Kaapse Platanna (Afrikaans)
Recommended citation format for this species text:
de Villiers AL, Tippett RM. Cape Platanna Xenopus gilli. BDI, Cape Town. Available online at http://thebdi.org/2021/12/15/cape-platanna-xenopus-gilli/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
de Villiers AL 2004 Xenopus gilli Cape Platanna. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Jennifer Sterne) in FrogMAPhere.
Find the Guttural Toad in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
GUTTURAL TOAD – Sclerophrys gutturalis
(Power, 1927)
Habitat
B. gutturalis inhabits various vegetation types in the Savanna, Grassland and Thicket biomes at altitudes ranging from sea-level to about 1800 m. In the east, the species is sometimes found in forest clearings and forest/grassland ecotones, while in the west it has a linear distribution along the wooded banks of the Gariep River.
These toads are opportunistic in their selection of sites for aestivation during winter. Quiescent toads have been found in termitaria and in the burrows of a large lizard, the Sungazer lizard Cordylus giganteus (Du Preez 1996).
Breeding takes place in open, shallow pools in rivers, farm dams, garden ponds, canals, ditches and borrow pits. During the day, the frogs take shelter under logs, rocks or other objects, in drain-pipes and gutters, in burrows, or in holes that they excavate in soft ground.
Behaviour
B. gutturalis is active from August to April, and occasionally during warm periods in winter. Breeding begins in small permanent water bodies, such as garden ponds, as soon as temperatures start to rise in spring, often before the first rains arrive. In areas where permanent water bodies do not exist, breeding is initiated by the first heavy spring rains. Dense choruses form, and the rolling, guttural snores of males can be heard from just after dark until well past midnight. Choruses may persist for several weeks, with a peak in late September to November, reviving after every rain shower until mid-February.
These toads usually call from partly concealed sites, although exposed sites are also used and calling males often seem oblivious to the presence of humans. Calling males exhibit site fidelity, with some individuals returning to the same site year after year. As many as 25 000 eggs, 1.4–1.5 mm diameter, are laid in two gelatinous strings, each 5 mm thick (Wager 1986). Strings of eggs are often twined around aquatic vegetation. Metamorphosis is complete after 5–6 weeks. As soon as the toadlets have developed front legs, they leave the water to hide on the bank until their tails are absorbed, and then start feeding on tiny insects. The maximum longevity, recorded in captivity, is seven years (Channing 2001).
At night they emerge from their shelters to prey on a wide range of insects, spiders and centipedes. In captivity, even lizards and other frogs have been recorded as prey items (Rose 1962). Individuals are often seen feeding on insects attracted to garden lights.
Adults are preyed on by the Black-necked Spitting Cobra Naja nigricollis, Common Night Adder Caususrhombeatus, Western Green Snake Philothamnus angolensis, Serrated Hinged Terrapin Pelusios sinuatus, and African Civet Viverra civetta (Channing 2001). The tadpoles are eaten by aquatic birds, fish and the Common Platanna Xenopus laevis.
B. gutturalis is relatively secure as it is widely distributed, locally abundant and highly adaptable to human settlement. It has been recorded in a number of national parks, provincial nature reserves and other protected areas. However, many individuals are killed by motor vehicles while crossing roads at night, especially during rain showers. On occasion, the toads are needlessly killed by people who are annoyed by their loud calls. The Eastern Cape lineage has a restricted distribution and further surveys are required in this area. Hybridization with B. rangeri occurs at scattered localities and may occur more frequently as a result of modern agricultural practices, but this does not seem to erode the differences between these species and is therefore not considered to be a threat.
B. gutturalis is distributed from western Uganda eastward to coastal Kenya and southward through Tanzania, Zambia, Malawi, Mozambique, Zimbabwe and Botswana, to South Africa (Tandy 1972). In the atlas region, its distribution is centred in the northeast, particularly in the KwaZulu-Natal, Mpumalanga, Gauteng, central Limpopo, eastern North West, and northern and eastern Free State provinces, and Swaziland. In the Northern Cape Province, its distribution extends westward along the Gariep (Orange) River as far as Goodhouse (2818CC). The species also occurs in Lesotho and northern Eastern Cape Province, but is absent from Western Cape Province except for an introduced population that became established on the Cape Peninsula (3418AB) in recent years.
Earlier workers, including Hewitt (1935, 1937), Poynton (1964), Guttman (1967) and Tandy (1972), recorded B. gutturalis as far south as Port St Johns (3129DA) in Eastern Cape Province. However, during the atlas period several more southerly populations were found, for example, at Hluleka Nature Reserve and surroundings (3129CC, CD), Cwebe Nature Reserve (3228BB), Amalinda Fish Station (3227DD) and Cintsa district (3228CC) near East London. This suggests that the species is expanding its range southward along the east coast. Alternatively, this may simply be the result of a more systematic survey than in the past.
Recent genetic studies of mitochondrial DNA have identified three geographically distinct lineages of B.gutturalis in the atlas region, with additional lineages further north (M.C. pers. obs). One lineage occurs near the Botswana border in the Gaborone district (2425DA), but the extent of this lineage south and west of this area is unknown. A second independent lineage occupies the central area of the range, including KwaZulu-Natal and Mpumulanga provinces and Swaziland. An introduced population in Mauritius, established in the 1940s, originated from this second lineage. The third lineage in the atlas region is restricted to the Eastern Cape populations, and meets the second lineage at Port St Johns. Toads in this southeastern population are smaller than those elsewhere, which led Hewitt (1935) to suggest that they represent a separate subspecies.
B. gutturalis and B. rangeri hybridize at several localities. The hybrids can be identified by aberrant morphology, genetic analysis, and by the fact that their advertisement calls are intermediate in structure to those of their parents. Hybrids have been found at Groenkloof (2528CA), Weza (3029DB) and Port St Johns (3129DA; Guttman 1967; Passmore 1972; M. Cherry in litt.), while an additional four hybrid populations were discovered during atlas surveys at Harding (3029DA), Pietermaritzburg (2930CB), Jamestown (3027CC) and Lubuyane Falls, Swaziland (2631AA).
The absence of records of B. gutturalis in parts of Northern Cape Province and Lesotho is probably due to inaccessibility of these areas rather than to real gaps in distribution. Apart from these limitations, the atlas data are reliable and reasonably comprehensive.
Distribution of Sclerophrys gutturalis. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gorrelskurwepadda (Afrikaans)
Recommended citation format for this species text:
du Preez LH, Weldon C, Cunningham M, Turner A, Tippett RM. Guttural Toad Sclerophrys gutturalis. BDI, Cape Town.
Available online at http://thebdi.org/2021/11/25/guttural-toad-sclerophrys-gutturalis/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
du Preez LH, Weldon C, Cunningham M, Turner A 2004 Sclerophrys gutturalis Guttural Toad. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Bart Wursten) in FrogMAPhere.
Find the Southern Foam Nest Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Rhacophoridae
SOUTHERN FOAM NEST FROG–Chiromantis xerampelina
Peters, 1854
Habitat
The species inhabits a variety of bushveld vegetation types in the Savanna Biome. Breeding usually takes place in temporary pans and vleis, but also occurs in more permanent water bodies such as dams and quarries. In the absence of trees and shrubs, nests may be attached to the sides of large rocks or man-made structures overhanging water, including bridges, culverts and bird hides.
Behaviour
In summer, these frogs are often seen perched on the branches of trees overhanging or near water, and their white, crusty nests, c.20 cm in diameter, are conspicuous around dams, pans and vleis and along river and stream courses.
In the winter months, they seek shelter under the bark of trees, in rock cracks and on the branches of shady evergreen trees far from the nearest water. They also move into buildings where they take up residence for weeks, or even months, on rafters, walls and windowsills, or behind bookcases and picture frames.
C. xerampelina is adapted in several ways to survive in an arid environment. It possesses a rough, dry skin, and conserves water by means of rectal water re-absorption and by excreting nitrogenous waste in the form of uric acid (Coe 1974). Inactive individuals may be found perched in exposed positions on branches of trees and shrubs where their colour becomes chalky white or pale grey to reflect light and heat.
The males gather at suitable nesting sites at night where they produce soft, discordant croaks and squeaks. They do not appear to be territorial, and two or more frogs close together, or even on top of each other, will call irregularly and independently.
The female leaves the water and climbs up to the nesting site where amplexus with one of the males takes place. Nest construction begins when the female releases an oviducal secretion from her cloaca and churns it into a white foam with her hind legs. Peripheral males take up positions on either side of the amplexing pair and attempt to position their cloacae adjacent to that of the female during bouts of oviposition. Thus the female’s eggs are fertilized by more than one male (Jennions et al. 1992). Neither the amplexing male nor the peripheral males participate in the construction of the foam nest.
Typical foam nest of Chiromantis xerampelina – Phongolo Nature Reserve, KwaZulu-Natal Photo by Ryan Tippett
The nest may take up to seven hours to complete, and nest construction is split into 2–4 sessions. Between sessions, the female leaves the nest site and returns to the water to rehydrate. At this time, the amplexing male may dismount and, on returning to the nest, the female may amplex with a different male (Jennions et al. 1992). Communal nests, involving two or more females and numerous males, are commonly formed. One such nest contained 50 males and 20 females (Passmore and Carruthers 1995). The female usually returns the following night and adds a second layer of foam (not eggs) to the top of the nest. Males seldom attempt amplexus on the second night; if they do, they soon release the female and leave (M.D. Jennions pers. comm.).
Jennions et al. (1992) recorded a mean clutch size of c.1200 eggs for single-female nests. Once the eggs hatch within the nest, the tadpoles rely on bubbles in the foam for oxygen (Seymour and Loveridge 1994). After 4–6 days, the wriggling tadpoles begin to move downward within the nest, sometimes in a wet squirming mass of several tadpoles or in ones and twos, until they reach the bottom (Wager 1986). It is thought that these movements and the accumulation of tadpoles at the bottom of the nest softens the crust, thereby enabling the tadpoles to drop into the water below where they complete their development. Egg development within a foam nest may serve to avoid or reduce predation in the early stages of tadpole development. However, in some cases, the water below the nest recedes, and the tadpoles drop onto the ground and perish.
Chiromantis xerampelina is preyed upon by arboreal snakes such as the Vine Snake Thelotornis capensis and Boomslang Dispholidus typus. The eggs are eaten by Samango Monkeys Cercopithecus mitis (Rödel et al. 2002) and the Greater Leaf-folding Frog Afrixalusfornasinii (Drewes and Altig 1996).
Status and Conservation
This widespread species is not threatened. Much of the natural habitat of C. xerampelina is used for game or stock farming and it occurs in numerous private and public protected areas.
Distribution
C. xerampelina is widely distributed in eastern and southern Africa. In the atlas region it ranges from Mafikeng (2525DC) in the North West Province, eastward through most of Limpopo Province and southward through the eastern lowveld of Mpumalanga and Swaziland to Empangeni (2831DD) in KwaZulu-Natal. In South Africa, the species occurs from near sea level in KwaZulu-Natal (Lambiris 1989a) to 1200 m in the former Transvaal (Jacobsen 1989).
Distribution of Chiromantis xerampelina. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Grootgrysskuimnespadda (Afrikaans)
Recommended citation format for this species text:
Boycott RC, Theron J, Tippett RM. Southern Foam Nest Frog Chiromantis xerampelina. BDI, Cape Town. Available online at http://thebdi.org/2021/11/24/southern-foam-nest-frog-chiromantis-xerampelina/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Boycott RC, Theron J 2004 Chiromantis xerampelina Southern Foam Nest Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Nick Evans) in FrogMAPhere.
Find the Pickersgill’s Reed Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Hyperoliidae
PICKERSGILL’S REED FROG – Hyperolius pickersgilli
Raw, 1982
Identification
H. pickersgilli is a small- to medium-sized Hyperolius species that reaches a snout–vent length of 22.3 mm in males and 28.5 mm in females (Raw 1982). The sexes are morphologically distinct.
Males and juveniles are light to dark brown, often with small black spots on the dorsum, and a dark-edged, white to silver, dorsolateral stripe extending from the tip of the snout, over the eye, to the groin. The lower surfaces of the limbs are yellow, while the abdomen and throat are yellowish to white. Adult males have a bright yellow gular sac.
As females mature, the dorsal colouration becomes a brilliant light to yellowish green and the dorsolateral stripe disappears, although a dark canthal streak from nostril to eye is sometimes present. The flanks are off-white to brownish white, and are distinctly demarcated from the dorsal colouration by an irregular margin. The ventrum is light yellow to white.
The males of H. argus, a sympatric species, possess a similar dark-edged, dorsolateral stripe, but the snout is less pointed and the thighs and concealed portions of the hands and feet are brown to orange or orange-red. H.pickersgilli females can be distinguished from H.tuberilinguis by the distinct border that separates the dorsal and lateral colouration in the former. H. pickersgilli is also substantially smaller than both H. tuberilinguis and H.argus (Raw 1982; Lambiris 1989a; Passmore and Carruthers 1995; Channing 2001).
Male H. pickersgilli produce unusually soft, cricket-like calls at irregular intervals. The call structure, with reference to published calls, is as follows: emphasized frequency: 3–3.3 kHz; duration: 0.1–0.7 s; number of pulses: 5–30; pulse rate: 50–100/s (Raw 1982; Passmore and Carruthers 1995). This variation may be partly due to a difference in the temperature at which the recordings were made (not indicated by the authors). A larger call sample, corrected to a common temperature, is required before a satisfactory description of the advertisement call can be given.
Habitat
H. pickersgilli inhabits Coastal Bushveld-Grassland, where it breeds in marshy areas containing dense stands of Saw Grass Cyperus immensus. The water at breeding sites is stagnant and rarely exceeds 50 cm in depth.
Behaviour
The behavioural ecology of H. pickersgilli in the non-breeding season is unknown. Calling takes place August–March, and froglets have been collected from late January to early March. Males call from elevated positions, well concealed in dense stands of sedges Cyperus spp.
A gelatinous mass of about 50 eggs is attached to vegetation, several centimetres above the water (Raw 1982). About one week later, tadpoles drop out of the egg mass into the water.
Raw (1982) noted that this species often occurs in sympatry with several other hyperoliids that also lay their eggs out of water, and speculated that this choice of oviposition site represents an adaptation to breeding in stagnant water with a low oxygen content.
H. pickersgilli has been listed as Rare (Branch 1988) and as Vulnerable (IUCN 2000). Its status was raised to Endangered in view of its small area of occupancy (<500 km2), severe fragmentation of its habitat, and evidence of a continuing decline in the area of occupancy, extent and quality of habitat, and number of locations (Harrison et al. 2001; this publication).
H. pickersgilli occurs in the following protected areas: Greater St Lucia Wetland Park, Umlalazi Game Reserve and Twinstreams-Mtunzini Natural Heritage Site. Outside of these areas it is protected by the KwaZulu-Natal Nature Conservation Ordinance, No. 15 of 1974, as amended. It is listed as a species of importance in KwaZulu-Natal (Goodman 2000).
Threats
The major threats to H. pickersgilli are ongoing habitat loss and fragmentation as a result of drainage for agricultural and urban development. The pollution of breeding sites in the vicinity of human settlements by DDT, which has been re-introduced to control malaria vectors, also poses a serious threat. Increase in human habitation and changes in land use are likely to have additional negative effects on the habitat, including the spread of alien vegetation.
Recommended conservation actions
A thorough distribution survey and a study of the life history and habitat requirements of this species are recommended. Known breeding habitats should be protected and monitored (Harrison et al. 2001).
Management recommendations include habitat management, limiting factor management, public education and monitoring programmes. This species often occurs in relatively small, stagnant ponds, which are more likely to be drained by landowners and municipalities than are larger wetlands. The public should be made aware of the importance of preserving these small pockets of breeding habitat.
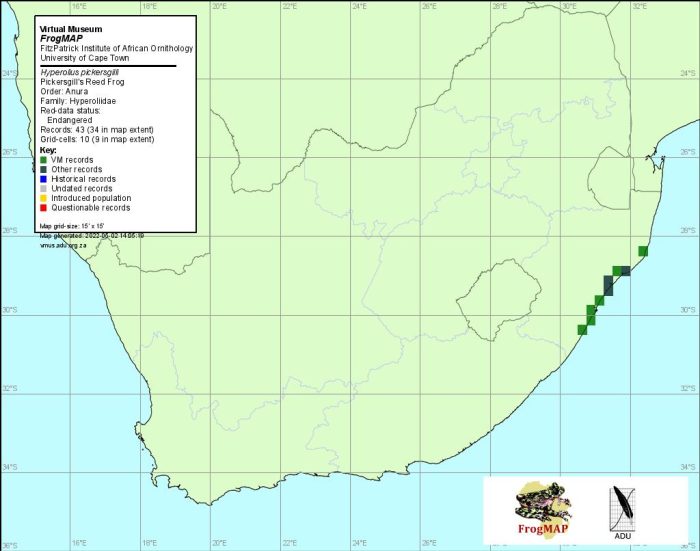
H. pickersgilli is endemic to KwaZulu-Natal, ranging from west of Kingsburgh (3030BB) in the south, along the coastal lowlands to St Lucia (2832AD) in the north. At least eight subpopulations (sensu IUCN criteria) are known (Harrison et al. 2001). The species is secretive, inconspicuous and easily overlooked: for example, at Twinstreams-Mtunzini Natural Heritage Site (2831DD), an area where frogs have been extensively studied for the past 25 years, this species escaped notice until fairly recently. Thus, although the area of suitable breeding habitat is relatively small, surveys may reveal additional populations.
It is interesting to note that this species is seldom found at the same breeding sites as H. marmoratus.
The atlas data may be regarded as reliable, but incomplete.
Distribution of Hyperolius pickersgilli. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Pickersgill se rietpadda (Afrikaans)
Recommended citation format for this species text:
Bishop PJ, Tippett RM. Pickersgill’s Reed Frog Hyperolius pickersgilli. BDI, Cape Town. Available online at http://thebdi.org/2021/11/24/pickersgills-reed-frog-hyperolius-pickersgilli/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Bishop Pj 2004 Hyperolius pickersgilli Pickersgill’s Reed Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Michael Adams) in FrogMAPhere.
Find the Red Toad in the FBIS database (Freshwater Biodiversity Information System) here.
Family Bufonidae
RED TOAD – Schismaderma carens
(Smith, 1848)
Identification
Schismaderma carens – Vaalkop Dam Nature Reserve, North West Photo by Lance Robinson
Habitat
This species inhabits a wide variety of vegetation types, primarily in the Savanna biome, but is also found in Grassland vegetation types, such as Rocky Highveld Grassland in Gauteng (Poynton and Broadley 1988; Lambiris 1989a). It breeds in deep, muddy pools or dams in these habitats.
Behaviour
When not breeding, S. carens has been found in caves, mine adits, burrows, and under stones, logs and piles of dead vegetation. It often enters houses, taking shelter in cupboards, plant pots, drawers and other unexpected places (Poynton and Broadley 1988; Jacobsen 1989; Lambiris 1989a). Individuals have even been found 2 m from the ground in trees. This toad seems to emerge earlier in spring and remains active later in autumn than most other summer-breeding species (Jacobsen 1989).
Breeding occurs in summer, usually at the peak of the rainy season. Calling has been recorded October–January in the Suikerbosrand Nature Reserve in Gauteng (Carruthers and Carruthers 1979). The low, booming call is produced while floating in water with limbs outstretched. Calling usually occurs at night but also on overcast, humid days. A large breeding aggregation at Hans Merensky Nature Reserve called throughout the night, laying their eggs in the early hours of the morning. By dawn, amplexing pairs were still present at the site but no egg-laying was observed (L.R.M. pers. obs.). Eggs are laid in double strings (cf. Stewart 1967) entwined around submerged vegetation. Estimates of egg numbers vary from about 2500 to 20 000 (Rose 1962; Stewart 1967; Passmore and Carruthers 1995; Channing 2001).
Tadpoles exhibit shoaling behaviour, forming dense clusters, 10–15 cm in diameter, that slowly move through the water, possibly aiding feeding by stirring up the substrate and creating a suspension of food particles (Wager 1965; Pienaar et al. 1976; Passmore and Carruthers 1995). This shoaling behaviour does not appear to deter predators as there are many records of fish, terrapins, birds and aquatic insects and their nymphs or larvae feeding avidly on these swarms (Pienaar et al. 1976; Channing 2001.). An interesting feature of the tadpole is the horseshoe-shaped fold of skin that extends backwards from behind the eyes to the middle of the body. This structure has a respiratory function, demonstrated by the fact that it is larger in tadpoles that are reared in polluted water with a low oxygen content. Under these conditions the tadpoles swim close to the surface (Charter and MacMurray 1939; Channing 2001).
Amplexus between S. carens and Bufo poweri has been observed in the wild (Power 1926a), but Blair (1972) was unable to induce artificial fertilization between these species. Given the great phylogenetic divergence between Schismaderma and Bufo, it seems unlikely that successful hybridization actually occurs and museum specimens identified as hybrids are probably anomalous individuals.
S. carens is not threatened. It occurs in numerous protected areas, including the Kruger National Park and adjacent private nature reserves, the Sandveld and Suikerbosrand nature reserves, and the Greater St Lucia World Heritage Area. Even in disturbed areas it adapts well to human habitation, occurring commonly in suburban gardens and homes. The ability of the tadpole to survive in polluted water favours its survival in human settlements, but adults and juveniles are often killed by traffic on roads.
Distribution
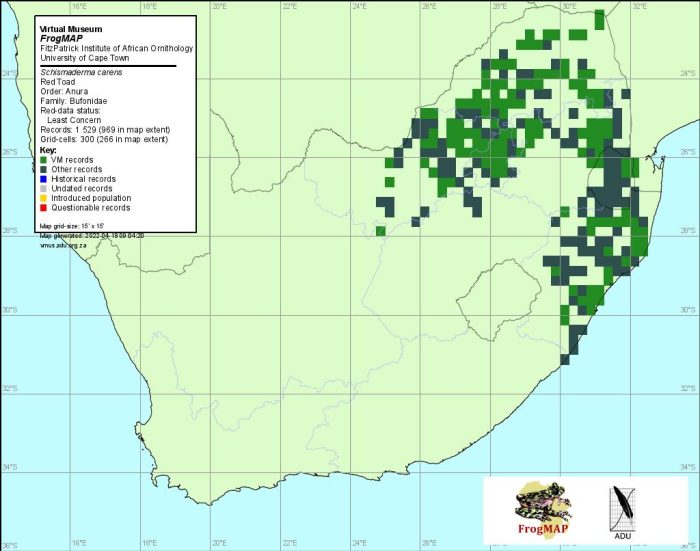
The Red Toad occurs from southeastern Democratic Republic of Congo and Tanzania, southward to Botswana, Zimbabwe and Mozambique. In the atlas region, S. carens is found from Vryburg (2624DC) in North West Province, eastward through the northern Free State, Gauteng, Limpopo Province, northern and eastern Mpumalanga and Swaziland, and southward throughout KwaZulu-Natal to Port Edward (3130AA). Within this area the species appears to be absent from the relatively cool highveld grasslands between Wakkerstroom and Dullstroom in Mpumalanga, and the relatively hot and dry Limpopo River valley.
The species cannot be confused with any other toad species and has a particularly characteristic call. The atlas data are reliable.
Distribution of Schismaderma carens. Taken from the FrogMAP database, April 2022.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Rooiskurwepadda (Afrikaans)
Recommended citation format for this species text:
Theron J, Minter LR, Tippett RM. Red Toad Schismaderma carens. BDI, Cape Town. Available online at http://thebdi.org/2021/11/24/red-toad-schismaderma-carens/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Theron J, Minter LR 2004 Schismaderma carens Red Toad. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Jean Hirons) in FrogMAPhere.
Find the Striped Stream Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pyxicephalidae
STRIPED STREAM FROG – Strongylopus fasciatus
(Smith, 1849)
Identification
Habitat
S. fasciatus inhabits a variety of vegetation types in the Forest, Fynbos, Thicket, Grassland and Savanna biomes. It occurs in well-watered areas with annual rainfall >500 mm, and it is rarely found far from permanent water (Greig et al. 1979). It ranges mainly through the summer-rainfall region, but extends into the winter-rainfall region in the southwest.
In montane grassland, these frogs seem to prefer grassy areas and reed beds along streams and rivers and around natural vleis. They are also found in well-vegetated man-made dams and ponds and along irrigation canals. They can tolerate disturbance and have been found in urban parks and gardens, and at dams surrounded by alien vegetation, in commercial forestry plantations.
Behaviour
Breeding takes place mainly in winter, and seems to be associated with a drop in temperature. The first calls are usually heard in mid- to late February, but there are records of calling as early as January (pers. obs.). In Swaziland, peak calling occurs in March, April and May, and calling activity ceases in November (pers obs.). Outside the winter months, sporadic calling may be triggered by a cold front moving through the subcontinent. In the KwaZulu-Natal midlands, strong choruses have been heard in midsummer (M. Burger pers. comm.).
At some breeding sites only a few calling males may be present, while at others, large choruses may form with calling males separated by only a few centimetres. Males call from the water’s edge or from elevated positions in reeds and grass.
The eggs are laid singly in shallow water on the edges of grassy pools, streams and man-made dams. They soon gather debris and become difficult to see. Although clutch size is not recorded for S. fasciatus, a clutch of 64 eggs was recorded for S. fuelleborni in Malawi (Stewart 1967), and another of 44 eggs for the closely related S.bonaespei of the Western Cape Province (Cunningham and Henderson 2000). This suggests that S. fasciatus does not lay large clutches of eggs. Tadpoles reach metamorphosis in 4–5 months (Wager 1986).
Status and Conservation
S. fasciatus is widespread and common and is not threatened. It often occurs in remote mountainous areas, but also survives in suburbia and other human-altered habitats, and appears to benefit from agricultural activities (Greig et al. 1979).
Distribution
S. fasciatus is found in the wetter, relatively temperate parts of the atlas region. Its range extends from Bonnievale (3320CC) in the Western Cape Province, eastward through the Eastern Cape Province to coastal KwaZulu-Natal, and inland to Lesotho and the Drakensberg escarpment of Mpumalanga and Limpopo provinces and Swaziland. It has a sporadic distribution in the high-altitude grasslands of the eastern Free State, Gauteng, North West and Limpopo provinces (Greig et al. 1979; Jacobsen 1989). The species frequently occurs at sea level, but many inland populations, for example in Swaziland, appear to be restricted to altitudes above 1100 m (pers. obs.).
North of the atlas region, S. fasciatus is found in highland areas of Zimbabwe, Zambia and Mozambique (Channing 2001).
This species is easily identified by its call. The atlas data are reliable and reasonably complete. The absence of recent records from summer-rainfall regions, such as Limpopo Province, may be due to inadequate surveying during the species’ winter breeding period.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gestreepde langtoonpadda (Afrikaans)
Recommended citation format for this species text:
Boycott RC, Tippett RM. Striped Stream Frog Strongylopus fasciatus. BDI, Cape Town. Available online at http://thebdi.org/2021/11/24/striped-stream-frog-strongylopus-fasciatus/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Boycott RC 2004 Strongylopus fasciatus Striped Stream Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Cornelia Rautenbach) in FrogMAPhere.
Find the Banded Stream Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pyxicephalidae
BANDED STREAM FROG – Strongylopus bonaespei
(Dubois, 1980)
Identification
Habitat
S. bonaespei typically inhabits Mountain Fynbos but is sometimes found on the margins of forest. It seldom occurs on steep slopes or in deep kloofs, preferring flatter, more open situations near streams. Annual rainfall in these areas is >500 mm.
Breeding takes place in shallow, seasonal, marshy areas and seepages that are well vegetated, usually with long grasses, stands of restios and sometimes ferns.
Although this species has been recorded from remnants of natural fynbos vegetation in a pine plantation (Greig et al. 1979), it occurs mainly in natural fynbos habitats and appears to be intolerant of disturbance.
Behaviour
S. bonaespei breeds mainly in winter but also calls in spring and late summer if conditions are suitable. In the southwestern Western Cape Province, breeding mostly takes place from May to early August. However, calling activity has been recorded up to mid-September, and intense calling has been heard as early as February (A.L. de V. pers. obs.).
Although dense breeding choruses have been heard in ideal conditions, especially early in the breeding season, the calls are usually scattered with calling males well spaced from one another. During a 45-minute period in the late afternoon, 20 active S. bonaespei adults were captured in a 70×35 m area of seepage (Greig et al. 1979). Males have been found calling from ground level at the base of overhanging vegetation near the water’s edge, and from 10–20 cm above ground level, spread-eagled between tall grass stems (Greig et al. 1979). Although these calling positions were noted at night, calling activity appears to be mainly diurnal. Observations at one site indicate peak calling activity around sunset with only occasional calls heard during the night (Cunningham and Henderson 2000).
The eggs are laid out of water on waterlogged earth or moss at the base of, for example, a restio or grass tussock, within about 5–20 cm of temporary pools or shallow runnels of water in seepage areas. The eggs are laid singly and may either be scattered, grouped in clusters, or deposited in rows of up to six or seven eggs. The clutch size is 39–104 eggs (Greig et al. 1979; Cunningham and Henderson 2000; A.L. de V. pers. obs.). Each egg is encased in a jelly capsule c.7 mm in diameter. The eggs develop into benthic tadpoles that complete their metamorphosis in water.
Status and Conservation
S. bonaespei is not threatened. It is relatively widely distributed and most of its habitat occurs in protected areas. These include the Cederberg, Grootwinterhoek, Boosmansbos wilderness areas, Limietberg, Hottentots Holland, Kogelberg, Grootvadersbosch, Maanschynkop, De Hoop, Outeniqua nature reserves, Tsitsikamma Forest, Agulhas and Cape Peninsula national parks, and others.
Threats to individual populations include invasive alien vegetation, tree plantations, wildflower farming, alteration of drainage patterns, too-frequent fires, building developments such as roads and dams, and urban encroachment in certain coastal areas. The most extensive of these threats is probably the spread of invasive alien vegetation that eliminates fynbos vegetation and reduces water resources. The control of invasive alien vegetation is the responsibility of the nature conservation authorities for the respective protected areas.
Distribution
S. bonaespei is endemic to the atlas region. Its distribution is restricted almost entirely to the mountain ranges of the Western Cape Province, extending marginally into the Eastern Cape Province. It is distributed from Hoogvertoon (3219AC) in the Cederberg in the north, to the southern foothills of the Soetanysberg (3419DD) in the south, and from the Cape Peninsula (3318CD, 3418AB, AD) in the west, to the Witelsbos Forest Reserve (3324CC) in the Tsitsikamma Mountains in the east.
Although S. bonaespei occurs mainly in mountainous areas, it has been found at a wide range of altitudes, from 10 m near the mouth of the Voëlgat River east of Hermanus, to 1670 m near Waaihoek Peak in the Hex River Mountains (Greig et al. 1979).
The atlas data are reliable and reasonably complete.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Gebande langtoonpadda (Afrikaans)
Recommended citation format for this species text:
de Villiers AL, Boycott RC, Tippett RM. Banded Stream Frog Strongylopus bonaespei. BDI, Cape Town. Available online at http://thebdi.org/2021/11/23/banded-stream-frog-strongylopus-bonaespei/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
de Villiers AL, Boycott RC 2004 Strongylopus bonaespei Banded Stream Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.
View the above photo record (by Johan Kritzinger) in FrogMAPhere.
Find the Namaqua Stream Frog in the FBIS database (Freshwater Biodiversity Information System) here.
Family Pyxicephalidae
NAMAQUA STREAM FROG – Strongylopus springbokensis
Channing, 1986
Identification
S. springbokensis resembles S. grayii in overall shape, but in profile it has a sharper snout. In S. springbokensis, the foot length is less than twice the head width, whereas in S. grayii it is more than twice the head width. The dorsum is yellowish brown with darker markings edged in dark brown. A narrow vertebral stripe is often present. Breeding males have small spines along the back of their legs and webbing develops along the margins of their fingers, making them broad and paddle-shaped (Channing 2001).
The advertisement call of S. springbokensis consists of a series of 2–7 notes with a total duration of 1.17 s (7 notes; Channing 2001). The call is easily distinguishable from the short clicking call of S. grayii. Aggression and male release calls are also produced.
Habitat
In relatively low-lying areas, S. springbokensis inhabits Upland Succulent Karoo vegetation in the Succulent Karoo Biome, while on the mountains it occurs in North-western Mountain Renosterveld of the Fynbos Biome. These areas receive annual rainfall of <60 mm. In this arid environment, these frogs are restricted to the proximity of springs, seeps, small permanent and non-permanent streams and artificial impoundments.
Behaviour
At the start of the winter rains, males begin calling from well-concealed call sites, usually some distance from water. The eggs are laid out of water, in the vicinity of the calling male. They are deposited under rocks or in rock crevices, tunnels in vegetation, or rodent burrows. Development is arrested after the tadpole’s eyes and tail are well formed. Tadpoles remain at this stage until rain floods the nest and then emerge from the egg capsules to continue their development in the water (Channing 1986).
Status and Conservation
Status
S. springbokensis was assigned to the category Data Deficient (Harrison et al. 2001) but was subsequently re-assessed and raised to Vulnerable (this publication). This decision was based on the results of additional atlas surveys that confirmed the limited and fragmented nature of the species’ distribution and the threats to its habitat. The species is distributed among at least four subpopulations (sensu IUCN criteria): Springbok, Richtersveld, Ghaamsberg and Kammiesberg. The habitat of each subpopulation is fragmented.
S. springbokensis occurs in Richtersveld National Park.
Threats
S. springbokensis is threatened by habitat loss which is expected to continue at the rate of <20% over the next 30 years. Increased grazing pressure, destruction of vegetation around breeding sites, siltation of streams, and pollution were identified as specific threats. A local threat in Springbok (the town for which this species is named) is the conversion of the type locality, an old reservoir in the town, to a duck pond, with the subsequent disappearance of this species from the site.
Recommended conservation actions
Additional distribution surveys and monitoring of populations at known breeding sites are recommended. Research aimed at identifying limiting factors should be conducted so that the habitat can be managed appropriately.
Distribution
S. springbokensis is endemic to the Northern Cape Province of South Africa. It is distributed from the Gariep (Orange) River valley southward through Namaqualand to Garies (3018CA), and as far east as Aggenys (2918BD). Altitude ranges from 200–1600 m. Distribution data are difficult to collect as the breeding sites are not obvious and are often inaccessible. The atlas data are reliable but probably incomplete.
Further Resources
Virtual Museum (FrogMAP > Search VM > By Scientific or Common Name)
More common names: Namakwa-langtoonpadda (Afrikaans)
Recommended citation format for this species text:
Channing A, Tippett RM. Namaqua Stream Frog Strongylopus springbokensis. BDI, Cape Town. Available online at http://thebdi.org/2021/11/23/namaqua-stream-frog-strongylopus-springbokensis/
Recommended citation format:
This species text has been updated and expanded from the text in the 2004 frog atlas. The reference to the text and the book are as follows:
Channing A 2004 Strongylopus springbokensis Namaqua Stream Frog. In Minter LR et al 2004.
Minter LR, Burger M, Harrison JA, Braack HH, Bishop PJ, Kloepfer D (eds) 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. Smithsonian Institution, Washington, and Avian Demography Unit, Cape Town.




























{kind=link}
{kind=link}